
干旱与复水对2种蟛蜞菊生长及生理生化特性的影响
2021-05-06 03:31朱宇林周兴文梁珠珠韦银霞
西北农林科技大学学报(自然科学版) 2021年4期
赵 英,吴 敏,邓 平,2,朱宇林,周兴文,梁珠珠,韦银霞
(1玉林师范学院,广西 玉林 537000;2西北农林科技大学 林学院,陕西 杨凌 712100;3广西大学 林学院,广西 南宁 530004)
干旱是影响植物分布和生长发育的主要环境因素,其危害程度居众多非生物胁迫之首[1]。干旱胁迫实质是植物缺水,不仅发生在干旱半干旱地区,而且在湿润和半湿润地区也常发生不同程度的阶段性干旱[2]。有研究表明,植物体内的抗氧化酶活性能在一定程度上反映其耐旱能力,抗氧化酶通过清除自由基,抑制脂质过氧化作用,从而保护植物细胞膜不受伤害[3];而细胞内渗透调节物质的增减,可以调节细胞液渗透势,维持细胞壁的弹性并保持膨压,提高植物细胞吸水或保水能力[4]。此外,有研究表明,解除干旱复水后,一些植物体内水分运输过程能否迅速恢复到胁迫前的状态[5],或在干旱复水后植物是否会出现生长加快、光合、蒸腾速率提高等补偿效应现象[6],也是评价植物耐旱性强弱的重要环节。因此,研究干旱与复水处理后,植物体内抗氧化酶活性和渗透调节物质含量的变化,可以了解植物在长期进化过程中的耐旱机制和策略。目前关于该方面的研究主要集中在农作物以及牧草[7-11]上,对于园林入侵植物的相关研究报道[12]较少。
三裂叶蟛蜞菊(Wedeliatrilobata(L.) Hitchc.),菊科(Composite),又称南美蟛蜞菊,多年生草本,原产热带美洲,是南方广大地区最具危害性的入侵杂草之一[13],严重威胁广西、广东、海南等地本地物种的定居及生长,破坏园林绿地生态系统的物种多样性[14]。蟛蜞菊(Wedeliachinensis(Osbeck. ) Merr.)与三裂叶蟛蜞菊同属,原产我国,在广西、广东、海南等地分布[15]。二者生活史特征相似,但蟛蜞菊生长较慢,对自然生态系统生物多样性无危害,其在南方园林绿地中常作地被植物使用,而三裂叶蟛蜞菊的入侵,改变了当地园林绿地生态系统的结构和功能。目前,相关研究多集中在对比二者对其他物种或群落的化感作用[16-19],入侵物种对根际土壤酶和土壤理化性质的影响[20-21],应对高温、酸雨和干旱等逆境时二者的表型可塑性、生物量分配、光合生理等[22-25]方面。而在干旱和复水处理下二者基本生理生化变化规律,揭示其耐旱能力与策略的研究[26]较少。
本研究通过自然干旱及复水处理,对比蟛蜞菊及三裂叶蟛蜞菊扦插苗生长、抗氧化酶、渗透调节物质、丙二醛和叶绿素含量的变化规律,探究二者对干旱胁迫环境的生理生态适应机理及其旱后恢复能力,以期为防止三裂叶蟛蜞菊入侵提供理论依据。
1 材料与方法
1.1 试验材料
于2018年5月20日清晨08:00,在广西壮族自治区玉林市玉林师范学院苗圃(22°64′N,110°14′E),选择生长健壮的蟛蜞菊(W.chinensis)和三裂叶蟛蜞菊(W.trilobata),采集长(35.13±0.85) cm(从匍匐茎顶芽向后测量)的匍匐茎各50枝,立刻用黑色塑料袋包裹,并将末端切口处浸置于水中,迅速带回实验室。分别将采集的试验材料剪成约5 cm带芽茎段(每个茎段保留2~3片叶子),在100 mg/kg ABT生根溶液中浸泡5 min,在玉林师范学院玻璃温室扦插苗床中进行软枝扦插;经4~5周培养后,选取生长健壮、长势基本一致的蟛蜞菊和三裂叶蟛蜞菊扦插苗各60株,移栽到塑料花盆(上口径×高×底径:13 cm×13 cm×11 cm)中,每盆2株,各30盆。培养基质为营养土、蛭石、塘泥,按照1∶1∶1的体积比混合,缓苗3周后开始干旱胁迫试验。为保证幼苗健康生长,育苗与缓苗期土壤含水量为(24.4±1.23)%。
1.2 试验设计
2018年7月2日在广西玉林师范学院玻璃温室大棚中进行干旱胁迫试验,棚内昼夜温度分别为28和20 ℃,白天光强为800 μmol/(m2·s),相对湿度保持在(60±2.5)%。蟛蜞菊和三裂叶蟛蜞菊分别设置对照组和干旱处理组,其中对照组20株(10盆),干旱处理组40株(共20盆,其中10盆用于复水试验)。整个干旱处理试验持续15 d,对照组土壤含水量保持在(24.4±1.23)%;干旱处理组采用自然干旱法;试验结束测定各处理幼苗指标。第16天,选取蟛蜞菊和三裂叶蟛蜞菊干旱处理另20株(10盆)幼苗进行3 d复水试验,使其土壤含水量达到(24.4±1.23)%,即达到对照组水平。试验第18天测定复水处理组(RT)相关指标。
1.3 试验方法
1.3.1 生长指标的测定 测量株高后,采用便携式叶面积仪(Yaxin-1241),测定蟛蜞菊和三裂叶蟛蜞菊茎干上、中、下部位各1片健康叶片的叶面积,每株3片叶子,每处理10株(5盆),取平均值。然后,收获整株植株的地上和地下器官并放置于牛皮纸袋中,放入烘箱于105 ℃下杀青10 min,再将温度调至 80 ℃烘干至恒质量,测定地上和地下部分干质量;计算总生物量和根冠比。
1.3.2 生理生化指标的测定 分别采集每个处理其余10株(5盆)蟛蜞菊和三裂叶蟛蜞菊茎干上、中、下部位健康新鲜叶片,均匀混合剪碎后称取0.2 g,生理生化指标测定均参考李合生[27]的方法,每个指标测定3次,取平均值。
测定指标包括叶绿素、丙二醛(malondialdehyde,MDA)、可溶性糖、可溶性蛋白含量及超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(APX)活性。
1.4 数据统计与分析
采用 SPSS 19.0软件对试验数据进行统计分析;采用单因素方差分析(one-way ANOVA)和 LSD检验蟛蜞菊和三裂叶蟛蜞菊各处理间的差异显著性(α=0.05),采用独立样本T检验分析相同处理蟛蜞菊和三裂叶蟛蜞菊种间的差异显著性(α=0.05)。采用Origin 9.0软件绘图。
2 结果与分析
2.1 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊生长的影响
由图1可以看出,干旱胁迫处理下,蟛蜞菊、三裂叶蟛蜞菊的株高和单叶面积均低于对照组,其中三裂叶蟛蜞菊的株高和单叶面积下降更显著(P<0.05),分别比对照组降低了28.41%和51.39%;且蟛蜞菊的株高和单叶面积均显著高于三裂叶蟛蜞菊。复水处理后,蟛蜞菊和三裂叶蟛蜞菊的株高、单叶面积均有不同程度提高,其中蟛蜞菊株高、单叶面积和三裂叶蟛蜞菊的株高均恢复到对照水平(P>0.05),而三裂叶蟛蜞菊的单叶面积则比对照组增加4.28%(P>0.05);株高和单叶面积在两物种间均无显著差异(P>0.05)。
由图1还可以看出,在干旱胁迫处理下,蟛蜞菊和三裂叶蟛蜞菊的总生物量分别比对照组显著降低31.55%和50.99%,根冠比则分别比对照组显著提高14.53%和38.59%;且蟛蜞菊总生物量比三裂叶蟛蜞菊高出41.01%(P<0.01),三裂叶蟛蜞菊根冠比则比蟛蜞菊高了31.00%(P<0.05)。在复水处理后,蟛蜞菊和三裂叶蟛蜞菊的总生物量均高于干旱处理组,但均显著低于对照组(P<0.05);反之,根冠比较干旱处理时降低,但均高于对照组,且三裂叶蟛蜞菊的根冠比较对照组显著提高14.97%(P<0.05);尽管复水处理后,三裂叶蟛蜞菊的总生物量和根冠比均高于蟛蜞菊,但均无显著差异(P>0.05)。
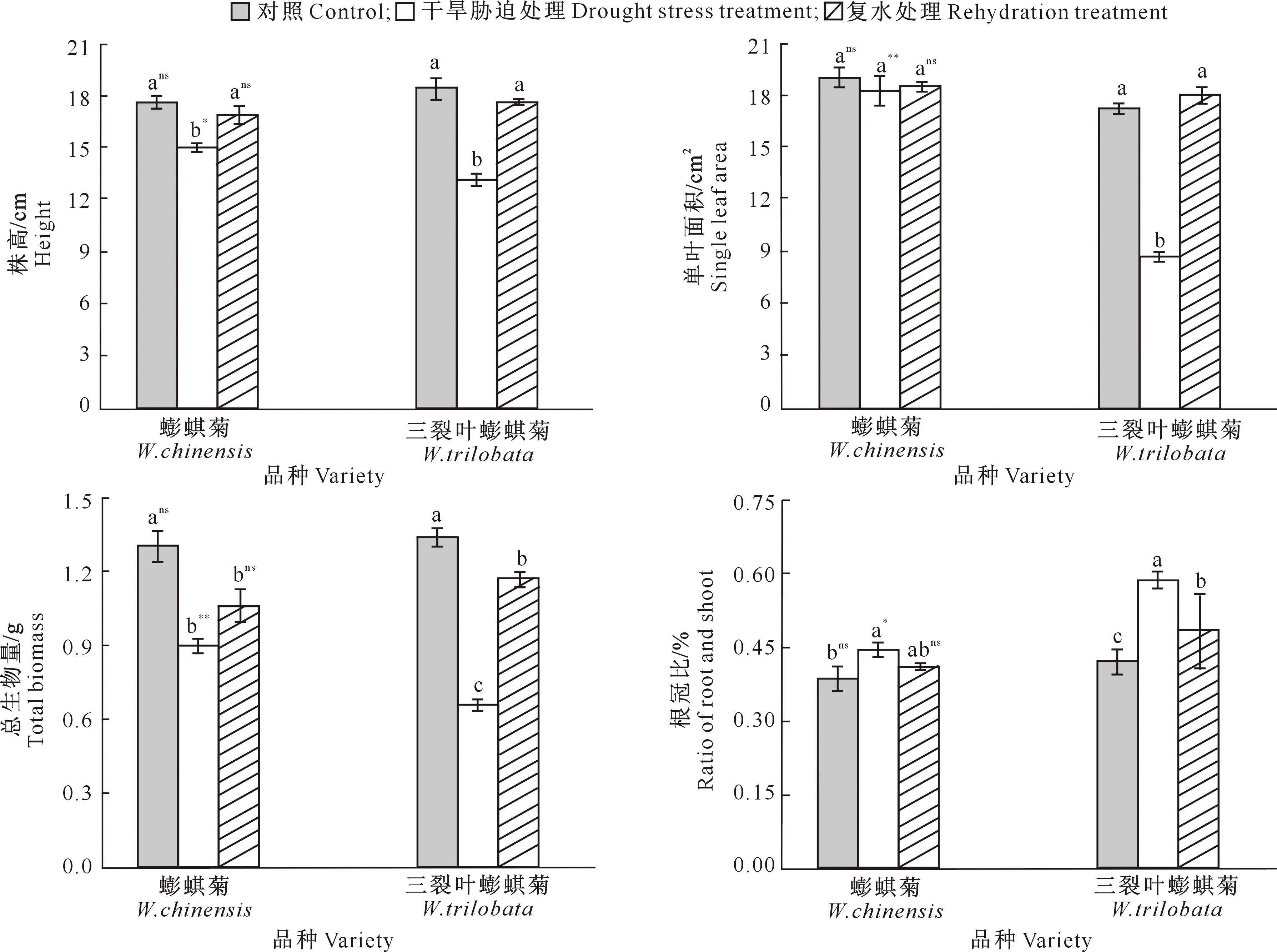
不同小写字母表示相同物种不同处理间差异显著(P<0.05); 在相同处理不同物种间,**表示差异极显著(P<0.01),*表示差异显著(P<0.05),ns表示差异不显著(P>0.05)。下同Different lowercase letters indicate significant difference in same species among different treatments at P<0.05.In same treatment between different species,** indicates significant difference at P<0.01;* indicates significant difference at P<0.05;ns indicates no significant correlation at P>0.05.The same below图1 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊生长的影响(n=10)Fig.1 Effects of drought and rehydration on growth of W.chinensis and W.trilobata (n=10)
2.2 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊叶绿素参数的影响
由图2可以看出,干旱胁迫处理对蟛蜞菊和三裂叶蟛蜞菊叶绿素a(Chla)含量均有显著影响(P<0.05),而对叶绿素b(Chlb)含量无显著影响(P>0.05);在干旱胁迫处理下,蟛蜞菊Chla含量比对照组增加了8.06%,而三裂叶蟛蜞菊Chla含量比对照组减少了33.36%,且前者极显著高于后者(P<0.01)。复水处理后,蟛蜞菊Chla含量减少并恢复到对照水平(P>0.05),三裂叶蟛蜞菊Chla含量则迅速增加(P<0.05),比蟛蜞菊Chla含量高出5.56%(P>0.05);复水处理对蟛蜞菊和三裂叶蟛蜞菊Chlb含量均无显著影响(P>0.05);且Chla和Chlb含量在种间均无显著差异(P>0.05)。
由图2可知,在干旱胁迫处理下,蟛蜞菊叶绿素a/b(Chla/b)和叶绿素总量(Chl(a+b))含量均显著高于对照组(P<0.05);三裂叶蟛蜞菊Chla/b和Chl(a+b)含量则均显著低于对照组(P<0.05),分别比对照组降低了31.69%和25.44%;且蟛蜞菊的Chla/b和Chl(a+b)含量分别比三裂叶蟛蜞菊极显著提高了66.18%和45.69%(P<0.01)。在复水处理后,尽管蟛蜞菊Chla/b和Chl(a+b)含量均比干旱处理时显著降低,但与对照组无显著差异(P>0.05);三裂叶蟛蜞菊Chla/b和Chl(a+b)均高于对照组,其中Chla/b比对照组增加了15.50%,差异达显著水平(P<0.05);且三裂叶蟛蜞菊Chla/b比蟛蜞菊高出13.23%,差异极显著(P<0.01)。

图2 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊叶绿素参数的影响(n=10)Fig.2 Effects of drought and rehydration on chlorophyll parameters of W.chinensis and W.trilobata (n=10)
2.3 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊MDA含量的影响
干旱与复水对蟛蜞菊和三裂叶蟛蜞菊丙二醛含量的影响见图3。
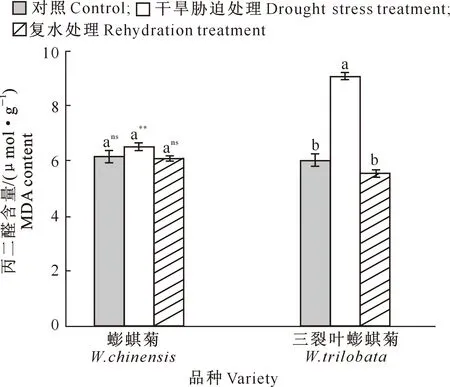
图3 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊丙二醛含量的影响(n=10)Fig.3 Effects of drought and rehydration on MDA content of W.chinensis and W.trilobata (n=10)
由图3可知,在干旱胁迫处理下,蟛蜞菊和三裂叶蟛蜞菊的丙二醛(MDA)含量均高于对照组,其中蟛蜞菊MDA含量与对照组之间无显著差异(P>0.05),三裂叶蟛蜞菊MDA含量比对照组显著增加了49.67%(P<0.05);且三裂叶蟛蜞菊MDA含量极显著高于蟛蜞菊(P<0.01)。复水处理后,蟛蜞菊和三裂叶蟛蜞菊MDA含量较干旱处理组均不同程度降低并恢复到对照水平(P>0.05),其中三裂叶蟛蜞菊MDA含量下降更快,比复水前显著降低了38.47%;且MDA含量在种间无显著差异(P>0.05)。
2.4 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊渗透调节物质含量的影响
由图4可以看出,在干旱胁迫处理下,蟛蜞菊和三裂叶蟛蜞菊的可溶性糖、可溶性蛋白含量均显著高于对照组(P<0.05),其中蟛蜞菊可溶性糖和可溶性蛋白含量分别比对照组增加了175.00%和40.15%,三裂叶蟛蜞菊分别比对照组增加86.29%和107.00%;且蟛蜞菊可溶性糖含量比三裂叶蟛蜞菊高出34.86%(P<0.05),而三裂叶蟛蜞菊可溶性蛋白含量比蟛蜞菊高出37.99%(P<0.01)。
在复水处理后,蟛蜞菊和三裂叶蟛蜞菊的可溶性糖、可溶性蛋白含量较复水前均不同程度降低,其中蟛蜞菊可溶性蛋白和三裂叶蟛蜞菊可溶性糖、可溶性蛋白含量下降较快并恢复到对照组水平(P>0.05),但复水后蟛蜞菊可溶性糖含量下降缓慢,仍显著高于对照组;蟛蜞菊可溶性糖和可溶性蛋白含量分别比三裂叶蟛蜞菊高出80.02%(P<0.01)和21.11%(P<0.05)。
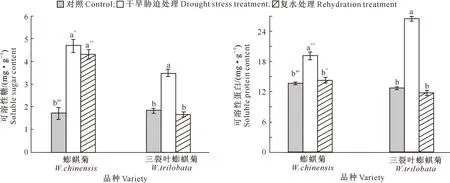
图4 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊渗透调节物质含量的影响 (n=10)Fig.4 Effects of drought and rehydration on contents of osmoregulation substances of W.chinensis and W.trilobata (n=10)
2.5 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊抗氧化保护酶活性的影响
干旱与复水对蟛蜞菊和三裂叶蟛蜞菊4种抗氧化保护酶(SOD、POD、CAT和APX)活性的影响见图5。
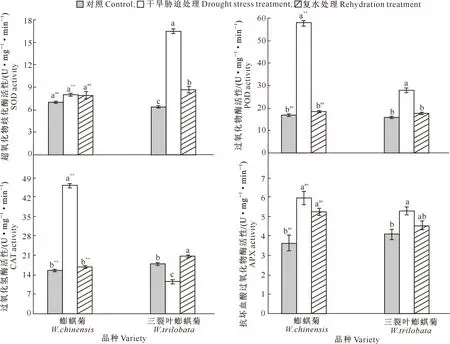
图5 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊抗氧化保护酶活性的影响(n=10)Fig.5 Effects of drought and rehydration on protective enzyme activity of W.chinensis and W.trilobata (n=10)
由图5可知,干旱胁迫处理下,蟛蜞菊4种抗氧化保护酶(SOD、POD、CAT、APX)活性均高于对照组,其中SOD活性与对照组无显著差异(P>0.05);三裂叶蟛蜞菊的SOD、POD和APX活性均显著高于对照组(P<0.05),分别比对照组增加156.00%,65.37%,63.08%,而CAT活性显著低于对照组(P<0.05);且三裂叶蟛蜞菊SOD活性极显著高于蟛蜞菊(P<0.01),而POD和CAT活性分别比蟛蜞菊低了51.92%和75.00%(P<0.01)。复水处理后,蟛蜞菊SOD、POD、CAT和APX活性均比复水前有不同程度下降,其中SOD、POD和CAT活性均恢复到对照水平(P>0.05),APX活性仍显著高于对照组43.12%(P<0.05);三裂叶蟛蜞菊SOD、POD、APX活性均低于复水前,其中POD和APX活性均恢复到对照水平(P>0.05),但SOD活性仍显著高于对照组35.16%(P<0.05),CAT活性上升,比对照组显著增加了15.07%(P<0.05);且在复水处理后,仅CAT活性在种间存在极显著差异(P<0.01),表现为三裂叶蟛蜞菊比蟛蜞菊高出24.37%。
3 结论与讨论
3.1 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊生长的影响
干旱是影响植物生长发育的主要限制因子,生长及各器官之间生物量分配比例的变化是植物应对环境胁迫的基本响应机制之一[28]。本研究发现,干旱胁迫抑制了蟛蜞菊和三裂叶蟛蜞菊的生长,单叶片面积减小,生物量显著下降,苗高生长受限;同时,干旱胁迫下蟛蜞菊和三裂叶蟛蜞菊根冠比增大,说明2种蟛蜞菊在一定程度上能通过增加根冠比调节地上和地下器官的生物量分配,以适应干旱环境。这与赵瑜琦等[29]对群众杨(Populus)扦插苗的研究结论相似。对比蟛蜞菊和三裂叶蟛蜞菊生长差异发现,尽管干旱处理下三裂叶蟛蜞菊根冠比增加更快,比蟛蜞菊高出31.00%,但并未显著遏制干旱胁迫对株高和叶面积的抑制趋势,甚至在干旱胁迫后期枯萎叶片数量明显高于蟛蜞菊,总生物量累积极显著低于蟛蜞菊。这与王宁等[30]对入侵植物节节麦(Aegilopstauschii)和何军等[31]对互花米草(Spartinaalterniflora)的研究结论相反,表明三裂叶蟛蜞菊通过增加根冠比未能达到规避干旱胁迫的效果,说明其耐旱性较蟛蜞菊弱。复水处理后,2种蟛蜞菊总生物量、株高、叶面积、根冠比均有所恢复,说明解除干旱胁迫后蟛蜞菊和三裂叶蟛蜞菊可以通过调节形态或生理生化反应恢复其生长[32],其中三裂叶蟛蜞菊各指标在复水后变化幅度较大,甚至出现超补偿效应,这可能是因为三裂叶蟛蜞菊叶面积和株高受干旱胁迫的抑制更严重,在复水后易产生超补偿效应,使其最大限度地快速恢复损失的营养器官,保证植株存活生长。这与肖凡等[33]对黄瓜(CucumissativusL.)幼苗的研究结论一致。
3.2 干旱与复水对蟛蜞菊和三裂叶蟛蜞菊生理代谢的影响
叶绿素含量是衡量植物光合生产潜力的一项重要生理指标,对植物光合速率、生物量生长等具有重要影响,也是衡量植物受环境胁迫程度的一项有效指标[34]。干旱胁迫后,蟛蜞菊主要通过提高Chla含量提高叶绿素总量,并在复水后保持较高的水平,这说明蟛蜞菊通过维持光合色素的稳定,提高其耐旱能力,这与冯慧芳等[35]对枫香(Liquidambarformosana)幼苗的研究结果一致。在干旱胁迫处理下,三裂叶蟛蜞菊则通过适当缩减叶绿素含量,减小叶片面积,降低蒸腾耗水,削弱干旱胁迫的伤害,复水后其叶绿素含量迅速恢复至正常水平,且Chla与总叶绿素含量与蟛蜞菊无显著差异,Chla/b显著高于蟛蜞菊,说明三裂叶蟛蜞菊叶绿素的合成并未受到干旱胁迫的根本性破坏,复水后通过自身调节可迅速进行旱后恢复。
渗透调节作用是植物在干旱胁迫下降低水势以及维持细胞膨压,从而抵御逆境胁迫的一种重要方式,可溶性糖、可溶性蛋白为有效的渗透调节物质[36]。在干旱环境下,蟛蜞菊和三裂叶蟛蜞菊的可溶性糖、可溶性蛋白含量均显著增加,说明2种蟛蜞菊在遭遇干旱胁迫时均可通过积累渗透调节物质,增大膨压吸收水分。复水后,三裂叶蟛蜞菊可溶性糖、可溶性蛋白含量迅速恢复到对照水平,表明解除干旱胁迫后三裂叶蟛蜞菊能够迅速恢复。蟛蜞菊复水后,可溶性糖含量虽略有降低但仍显著高于对照,且蟛蜞菊可溶性糖和可溶性蛋白含量均显著高于三裂叶蟛蜞菊,这与王好运等[37]对马尾松(Pinusmassoniana)的研究结论相反,可能是由于复水时间较短,蟛蜞菊细胞还保留大量的渗透调节物质,因此可以保持较高膨压吸收水分。
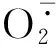
综上所述,蟛蜞菊可以通过增加叶绿素含量,积累渗透调节物质,抗氧化保护酶系统的协调作用,降低丙二醛含量,从而抵御干旱环境,保证植株的生长发育;复水后其叶绿素、渗透调节物质含量以及抗氧化保护酶活性均下降,且大部分能恢复至正常水平,说明蟛蜞菊对干旱胁迫造成的损伤具有较高的修复能力。尽管外来入侵种三裂叶蟛蜞菊也可通过调节抗氧化保护酶系统、渗透调节物质、增加根冠比等协同作用抵抗干旱,但在干旱处理期间,其株高生长缓慢,茎干萎蔫,叶片枯黄且叶面积缩减明显,说明其耐旱性较蟛蜞菊差,复水后其恢复能力较好。因此,建议今后在园林绿地管理中,可适当降低绿地土壤的含水率,从而抑制三裂叶蟛蜞菊的生长。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
林业调查规划(2022年3期)2022-06-13
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
少儿科学周刊·少年版(2021年17期)2021-01-17
阅读(科学探秘)(2020年8期)2020-11-06
安徽农学通报(2020年7期)2020-05-26
绿色科技(2019年2期)2019-05-21
江苏农业科学(2017年19期)2017-11-22
