
不同温度调控下西红花花芽分化进程及内源激素动态变化
2021-05-06 03:31李青竹许俊旭张永春
西北农林科技大学学报(自然科学版) 2021年4期
王 桢,李 心,李青竹,许俊旭,杨 贞,张永春
(上海市农业科学院 林木果树研究所,上海201403)
植物花芽分化和开花是一个复杂的生理生化和形态变化过程,是由营养生长向生殖生长(开花)的转变,与信号网络的不同排列组合紧密相关,这些信号包括外界的刺激如温度和光照等[1-2]。在许多植物中,开花时间受到光照长短(光周期)和温度的精确调控,特别是温度已经被证实能极大地影响开花时间。在模式植物拟南芥中,高温(27 ℃)处理能够促进拟南芥开花[3],而低温(16 ℃)下生长的拟南芥则表现为晚花,长日照、低温(16 ℃)生长条件下的拟南芥比23 ℃(对照)处理表现出开花延迟[4]。球根类植物由于其特殊的形态结构,在生理特性、养分运输、环境因子响应等方面均具有一定的特异性,有关球根花卉成花诱导及开花机制的研究中,针对百合[5-6]、鸢尾[7]、水仙[8-9]、郁金香[10-11]等的研究相对较多,已经深入到分子和蛋白水平。水仙(Narcissustazetta)的花芽起始和分化发生在夏季的高温休眠阶段,相比低温(12 ℃)环境,高温(25~30 ℃)能显著促进水仙开花[9];同时也有研究发现,环境温度对郁金香(Tulipagesneriana)成花转变具有重要的影响,冬季休眠后,相比较低的温度(8~9 ℃),较高的环境温度(18 ℃)能促进其成花[11]。
西红花(CrocussativusL.)为鸢尾科西红花属多年生球根草本,又名藏红花、番红花,原产于伊朗、西班牙和希腊等地中海国家,目前以伊朗的栽培面积最大。其干燥柱头可入药,是一味名贵中药材,也是世界上最昂贵的香料作物之一。20世纪80年代初,上海市药材公司先后从日本、德国引进西红花种球资源进行栽培研究,目前已发展成为上海的地道中药材。现在在上海、安徽、浙江、江苏等很多地区已经形成了一定面积的栽培规模。
西红花普遍采用“两段法”栽培模式,由于其不能进行有性繁殖,生产上多采用分球繁殖方式,每年11月至次年5月在大田繁育新的子球茎替代母球茎,6-11月是室内储藏-开花阶段,其间5-6月为休眠期,7-8月进行叶芽、花芽的分化和叶片、花器官的形成,9月顶芽萌发伸长,花期一般从10月底始花持续到11月中旬。西红花在室内储藏-开花的特性为通过控制温度等环境条件来调控花期提供了可能性。笔者前期对西红花子球茎膨大过程的形态学变化过程和生理指标的变化进行了观测[12],为通过调控室内环境条件来调控花芽分化和开花奠定了前期基础。
有研究发现,西红花开花前需要经历炎热的夏季高温阶段,较高的环境温度对打破西红花芽休眠和开花起始是必需的[13],前期西红花种球在23~27 ℃下处理50 d以上,再转移至17 ℃下开花量最大[14],25 ℃处理105 d后开花最早[15],此外前期2 ℃冷藏处理会延迟开花[16]。近几年,国内关于西红花花芽分化过程及生理指标变化有少量报道[17-19],基本明确了花芽分化的起始时间,但是花芽分化和发育过程中主要营养物质和激素变化存在异同,关于温度对其花芽分化、发育和开花阶段的影响也有待探讨。
花芽分化作为西红花球茎从营养生长向生殖生长转变的关键时期,其早晚和有无直接影响后续的开花率、开花时间及柱头产量等。本研究以西红花顶芽(花芽)作为研究对象,对不同温度处理条件下西红花顶芽的形态学变化过程和内源激素等进行观察测定,明确西红花花芽分化及后续开花阶段需要的适宜温度条件,以期为室内环境调控其开花提供理论依据,为其花期调控、生产实践等方面提供指导。
1 材料与方法
1.1 试验材料
试验材料为西红花(CrocussativusL.)种球,种植于上海市农业科学院林果所青浦白鹤花卉基地。2018年5月上旬收获子球茎,晾干后储藏于阴凉通风干燥处,挑选单球质量16~20 g的健康种球作为供试材料。
1.2 试验设计
试验于2018年5月至11月在上海市农业科学院奉浦院区进行。(1)选取1 000颗球茎随机分成3组,作为3次重复。从06-15开始,将其置于自然温度,湿度60%~80%,光照以暗为主,通风良好的室内培育房中,顶芽向上排放于培养架上,并测量顶芽长度。前期预试验结果表明,6月西红花处于休眠期,顶芽无肉眼可见的变化。因此,于07-05开始,每周测量顶芽长度,并以3 d为间隔取样,每组随机采集生长一致的西红花球茎顶芽各3个,于福尔马林-醋酸-乙醇(Formalin-acetic acid-alcohol,FAA)固定液中固定,用于石蜡切片,观察花芽分化形态变化过程。
(2)选取2 000颗球茎随机分成4大组,每组再随机分成3小组作为3次重复。06-15,分别将每组球茎置于15,20,25和30 ℃恒温培养箱中,湿度60%~80%,6-9月光照以暗为主,10月上旬开始16 h/8 h光暗条件,光强为100 μmol/(m2·s),测量顶芽长度,同时选取5个西红花种球的顶芽,液氮处理后冻存于-80 ℃冰箱中。于07-05开始,每周测量不同处理顶芽长度,同时每小组选取3个西红花种球顶芽,用FAA固定液固定,石蜡切片用于花芽分化的显微观测(顶芽长度小于7 mm),或者直接观察(顶芽长度大于7 mm),共进行10次;于07-05开始,每周每小组另选取5个西红花种球的顶芽,液氮处理后冻存于-80 ℃冰箱中备用,共采样8次,根据花芽分化观测结果选择相应采样点样品用于激素含量测定。其中,花芽分化完成后,将25 ℃恒温处理下的球茎于10月上旬转移至15 ℃恒温培养箱中至开花,统计不同温度处理下花芽分化的球茎比例。
1.3 试验方法
1.3.1 顶芽形态发育过程观察 取新鲜球茎的顶芽部位,于FAA固定液中固定24 h,然后进行50%,70%,85%,95%,100%乙醇梯度脱水,再于二甲苯中透明,包埋于石蜡中。用Leica显微切片机切成厚度8~10 μm的切片,脱蜡透明,爱氏苏木精染色,Olympus光学显微镜下观察并拍照[20]。另外,顶芽长度大于7 mm后,可直接剥离外层的白色叶鞘和包裹花器官的半透明薄膜结构,对叶片、花器官等进行观察拍照。
1.3.2 激素含量测定 根据不同温度下花芽分化进程显微镜观察结果,选择15 ℃(无花芽分化,叶芽)和25 ℃(有花芽分化,花芽)条件下,对7次采集的顶芽样品进行脱落酸(ABA)、生长素(IAA)和GA3含量的测定。激素含量测定采用改良高效液相色谱(HPLC)方法[21],以鲜质量计,各时间点采集的顶芽样品设2个生物学重复。
1.3.3 数据处理 试验数据采用Excel软件进行数据处理和制表,利用SPSS软件进行数据分析。
2 结果与分析
2.1 自然温度下西红花花芽分化进程的形态学观察
通过对不同时期的西红花顶芽石蜡切片进行观测发现,西红花顶芽从营养生长向生殖生长转变,至形成完整的花器官到最终开花,需要经历一系列的形态发育过程。根据其花芽分化的先后顺序与形态特征,可将其整个分化过程大致分为5个阶段。
2.1.1 未分化期 7月中旬,顶芽均未进入分化期,此时生长锥呈半圆球状,宽度大于高度。随着生长锥的进一步生长,由生长锥顶端分生组织侧面的表层细胞分裂而来的叶原基不断形成,呈小指状突起,向两侧扩展,包围生长锥(图1-A)。
2.1.2 分化初期 最早出现在7月下旬,一直持续到8月初,顶芽生长锥陆续开始由营养生长转变为生殖生长状态。生长锥顶部逐渐变圆滑肥大,向上隆起,呈半球形(图1-B)。
2.1.3 花原基分化期 8月上旬,大部分西红花球茎顶芽生长点可明显观察到2个形状相似、大小各异的突起。生长锥继续伸长增长,在隆起的半球形生长锥上分化出几个突起,即为花原基(图1-C)。08-02顶芽长度为(0.50±0.09) mm(图2)。

(A) 7月中旬,未分化期,顶芽侧翼长出叶原基,被低出叶包裹;(B) 7月底,分化初期,顶芽从营养生长转变为生殖生长;(C) 8月上旬,花原基分化;(D) 8月中旬,花被片原基和雄蕊原基分化期;(E) 8月下旬,产生明显的花被片和雄蕊;(F) 9月上旬,雌蕊原基分化,产生明显的雌蕊和心室。c.低出叶;l.叶片;b.苞片;fp.花原基;t.花被片;st.雄蕊;g.雌蕊;ve.心室;图4同(A) In mid-July,vegetative stage:shoot apex with leaf primordia initiated at the flank of the meristem,wrapped in cataphylls;(B) At the end of July, initial differentiation stage:shoot apex was transferred from vegetative to reproductive stage;(C) In early August,the flower primordium initiated;(D) In mid-August,perianth primordium and stamen primordium produced;(E) In late August,tepals and stamens produced; (F) In early September,gynoecium and ventricles produced.c.Cataphyll;l.Leaf;b.Bract;fp.Floral primordium;t.Tepal;st.Stamen;g.Gynoecium;ve.Ventricle;The same for Fig.4图1 自然温度下西红花花芽分化进程Fig.1 Process of floral bud differentiation of saffron under natural temperature
2.1.4 花被原基及雄蕊原基分化期 进入花原基分化期后,生长锥进一步升高,于中央及基部周边隆起,且逐渐分离,两侧各出现一片凸起,即为外轮花被原基。西红花有3枚外轮花被原基,石蜡切片显示为2枚大小不等的侧面突起;接着在外轮花被原基内侧又逐渐形成1对相对较小的凸起,与外轮花被原基对生,即为内轮花被原基(图1-D)。花被原基形成后,在其内侧基部出现突起,经过快速分裂、分化形成雄蕊原基。西红花有3枚雄蕊,石蜡切片可明显观测到2枚(图1-E)。从花被分化过渡到雄蕊分化,进程很快,一般8月中旬西红花球茎顶芽基本完成花被原基及雄蕊原基的分化。
2.1.5 雌蕊原基分化期 继雄蕊原基分化完成后,花芽原基的顶端开始凹陷,并逐渐发育形成3个突起,向上生长,即雌蕊原基。一般8月底至9月上旬西红花球茎顶芽基本完成雌蕊原基的分化,逐渐伸长长大,并伴随心室和胚珠的形成与发育(图1-F)。09-10顶芽长度为(7.22±1.48) mm(图2)。

图2 自然温度下西红花顶芽长度的变化Fig.2 Changes of shoot length of saffron under natural temperatures
2.2 不同温度下西红花顶芽(花芽)分化进程的比较
不同恒温(15,20,25和30 ℃)处理的西红花顶芽表现出花芽分化和生长发育的巨大差异。如表1所示,15 ℃恒温处理下花芽分化的球茎比例为0,且顶芽中只有叶片生长;20 ℃恒温处理下,约有55%的球茎顶芽能进行花芽分化并开花;而在25 ℃恒温处理和自然温度下,该比例达到95%以上;但是持续30 ℃温度下,在处理时间内未观测到顶芽萌发伸长现象,后续转移至20~25 ℃适宜温度后,只观测到顶芽萌发和叶片生长,未见花芽分化和开花。

表1 不同温度下西红花花芽分化时间及比例Table 1 Duration and ratio of floral bud differentiation in saffron under different temperatures
同时,在自然温度下,花芽分化的起始时间在7月底左右,一直持续到9月上旬,持续时间45~50 d。而在25 ℃恒温下,花芽分化的起始时间提前至7月中旬左右,持续到8月中下旬,持续时间30~40 d。说明温度能显著影响西红花顶芽的花芽分化与否和快慢,适宜的温度能使成花转变阶段提前,并且花芽分化持续时间缩短。
如图3所示,从6月中旬开始,15 ℃下西红花顶芽萌发最早,特别是从7月下旬开始,伸长生长速度比20和25 ℃处理球茎顶芽快,可以推测一定温度范围内,温度高低与顶芽伸长生长速度呈反比。09-03,15和20 ℃处理顶芽长度分别为(44.39±2.09)和(40.80±1.01) mm,25 ℃处理顶芽长度仅为(7.98±0.79) mm。
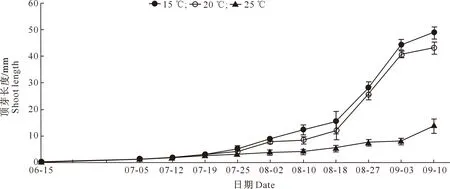
图3 不同温度下西红花顶芽长度的变化Fig.3 Changes of shoot length in saffron under different temperatures
于7月初开始,每周采集15和25 ℃顶芽进行显微和形态学观测。如图4所示,在15 ℃条件下,顶芽的生长锥保持营养生长状态,从生长锥侧翼产生叶原基和叶片,随着顶芽的萌发伸长,叶片逐渐伸长,颜色逐渐从淡黄色变为黄绿色和绿色,叶片数最终维持在7~11片(图4-I1)。在25 ℃条件下,最早从7月中旬开始,生长锥向上隆起,有向生殖生长转变的趋势(图4-B2);07-19,顶芽生长锥明显向上隆起,出现不规则突起,进入花芽分化初期(图4-C2);07-25,顶芽长度达到(3.12±0.48) mm,明显可见2个完整的花原基,有苞片、花瓣、雄蕊等(图4-D2);08-18,剥开管状低出叶后,可见叶片和花被、雄蕊(图4-G2),剥除雄蕊后,可见几乎无色的雌蕊;09-10,顶芽长度达到(13.84±2.87) mm,剥开管状低出叶后,可见花瓣和黄色的花药组织,以及环绕的叶片(图4-J2)。

(A1-D1)从6月中旬开始,15 ℃西红花顶芽的形态学观测;(E1-J1)剥离顶芽外包裹的低出叶(叶鞘)后,明显可见叶片。(A2-F2)从6月中旬开始,25 ℃西红花顶芽的形态学观测;(G2)8月18日,剥离顶芽外包裹的低出叶(叶鞘)、苞片和雄蕊后,可见明显的叶片、花被和柱头;(H2-J2)剥离顶芽外包裹的低出叶(叶鞘)和苞片后,可见明显的叶片、花被和雄蕊,雌蕊位于雄蕊内轮(A1-D1) From mid-June,morphological observation of shoot apex of saffron under 15 ℃;(E1-J1) Leaves were obvious after removing sheath of cataphylls.(A2-F2) From mid-June,morphological observation of shoot apex of saffron under 25 ℃;(G2) On August 18th,leaves,tepals and stigma are shown with the sheath of cataphylls,bract and stamen removed;(H2-J2) Leaves,tepals and stamen are shown with the sheath of cataphylls and bract removed,pistils are inside the stamen图4 不同恒温处理下西红花顶芽形态学变化过程Fig.4 Process of floral bud differentiation in saffron under different constant temperatures
2.3 不同温度下顶芽(花芽)分化过程中内源激素含量的变化
基于不同恒温处理下的花芽分化进程观察结果发现,从6月中旬开始至07-05第1次采样,顶芽均保持营养生长状态,同时25 ℃处理下的球茎于7月中旬开始陆续进入花芽分化初期。因此对07-05至08-27采样期间,15和25 ℃温度下的顶芽(花芽)进行内源激素含量测定。
如图5-A所示,15 ℃处理脱落酸(abscisic acid,ABA)含量较25 ℃处理要高,与处理前相比,整体均呈下降趋势;但25 ℃处理呈现“上升-下降”趋势。25 ℃处理下,ABA含量在07-12,即花芽分化初期最低,为(1.11±0.03) μg/g,说明ABA对花芽萌发的抑制作用减弱,花芽分化开始之后ABA含量逐渐升高,峰值出现在08-02,即花被片、雄蕊发育期,之后ABA含量急剧降低。15 ℃处理下,ABA含量整体呈下降趋势,07-12顶芽开始萌动并产生叶片,此时ABA含量为(11.88±0.62) μg/g,随着顶芽和叶片的进一步萌发伸长,ABA含量逐渐降低。说明低水平的ABA有利于西红花花芽向生殖生长转变,形态分化开始后,ABA含量略有升高,花器官、叶片形成后,又趋于下降。
如图5-B所示,15 ℃处理生长素(indole-3-acetic acid,IAA)含量较25 ℃处理要高,与处理前相比,整体均呈下降趋势。25 ℃处理下,花芽分化过程中呈现上升趋势,与15 ℃趋势相似,07-12,即花芽分化初期,IAA含量最低为(0.91±0.09) μg/g,之后含量升高,在07-25花被、雄蕊分化期达到一个峰值;08-18,此时顶芽进一步发育,肉眼可见完整的各轮花器官和叶片,IAA含量达到(3.28±0.27) μg/g,是分化初期的3.6倍。说明低水平的IAA有利于西红花从营养生长向生殖生长的转变,较高水平的IAA有利于西红花花芽分化的进行和花器官发育。
如图5-C所示,15 ℃处理GA3含量较25 ℃处理要低,整体分别呈现“下降-上升”和“上升-下降”的相反的变化趋势。15 ℃处理下,顶芽萌动前期,GA3含量逐渐下降,随着叶片的伸长发育,从08-02开始上升。25 ℃处理下,07-12,即花芽分化初期,GA3含量较处理前有所上升,为(8.09±0.14) μg/g,之后逐渐升高,于07-25花被、雄蕊分化期达到峰值,与IAA含量变化相似,之后GA3含量略有降低;08-18顶芽进一步发育,GA3含量达到(9.58±0.39) μg/g,仍然高于花芽分化初期(07-12)。说明GA3的积累有利于西红花花芽向生殖生长转变,且在分化过程中含量逐渐升高,说明较高水平的GA3有利于花器官的分化和发育。

图5 不同恒温处理下西红花顶芽分化过程中内源激素含量的变化Fig.5 Changes of endogenous hormones during shoot apex differentiation in saffron under constant temperatures
2.4 不同温度下西红花顶芽(花芽)分化过程中内源激素含量比值的变化
如图6所示,在西红花顶芽分化(15 ℃)之前,ABA/GA3和IAA/GA3均呈现下降趋势,但从07-12开始,叶片开始形成,两者变化出现差异。ABA/GA3持续降低,且在后期随着叶片伸长,降低幅度加大;而IAA/GA3开始逐渐升高,07-25出现一个峰值后,又开始降低。ABA/IAA在顶芽分化以前升高,07-12达到峰值,顶芽分化开始后,持续降低。整体而言,较高的ABA/GA3和ABA/IAA有助于维持顶芽的营养生长状态和叶原基的萌发,随着叶片的伸长生长,比值均逐渐下降。

图6 不同恒温处理下西红花顶芽分化过程中内源激素含量比值的变化Fig.6 Changes in ratios of endogenous hormones during shoot apex differentiation in saffron under constant temperature
如图6所示,在西红花花芽分化过程(25 ℃)中,ABA/GA3和IAA/GA3变化幅度不大,一直处于较平稳的状态,但是相比6月中旬和07-05花芽未分化期的高比值,ABA/GA3和IAA/GA3都有急剧降低的过程,并在07-12花芽分化初期有低峰值。比对15 ℃处理下,只有叶片萌发生长的顶芽中内源激素比值变化,说明较低的ABA/GA3和IAA/GA3有助于西红花顶芽向生殖生长转变。由于ABA和IAA含量都在花芽分化初期降低,分化过程中略有升高,整体变化趋势说明较低的ABA/IAA有助于顶芽向生殖生长阶段的转变,花器官形成后比值下降。
3 讨 论
3.1 温度对西红花花芽分化和生长发育的调控作用
本研究对西红花顶芽进行解剖结构观测后发现,自然温度下,7月底开始,顶芽从营养生长向生殖生长转变,主要经历花原基分化期、花被原基及雄蕊原基分化期、雌蕊原基分化期等3个关键的时期,整个分化过程历时45~50 d,属于向心型发育。同时,花芽分化过程中会出现分化不同步的现象,可能是由于群体较大和个体差异,相同时间内会出现不同分化时期的重叠,但是某个时间段会有分化高峰期。9月上旬,几乎所有球茎的顶芽已经形成完整的花器官,之后顶芽进一步伸长,进入顶芽(花芽)发育期。
西红花一般采用“两段法”栽培模式,为通过控制室内条件来调控花期提供可能性。有文献报道西红花在25 ℃处理105 d后,转移至17 ℃下开花最早,同时,西红花种球前期在23~27 ℃中处理50 d以后,再转移至17 ℃下开花量最大[15,22]。
本研究通过不同恒定温度(15,20,25和30 ℃)处理,观测了其顶芽的萌发时间、生长速度和花芽分化的比例等。结果表明,从6月中旬处理开始,15 ℃下顶芽萌发最早,生长速率最快,20和25 ℃下次之,但是30 ℃下,顶芽一直未萌发,说明持续的高温条件会抑制顶芽的花芽分化和萌发。因此,在一定温度范围(15~25 ℃)内,温度高低与顶芽伸长生长速度呈反比。同时,花芽分化的观测结果表明,15 ℃下球茎顶芽均无花芽分化,而在20和25 ℃条件下,花芽分化的球茎比例分别为55%和96%,说明西红花花芽分化所需温度不能低于20 ℃,也不能高于30 ℃。通过观测25 ℃下西红花花芽分化的进程和持续时间可以发现,其花芽分化的时间为7月中旬至8月中旬,共30~40 d;而在自然温度下的花芽分化进程,从7月下旬至9月上旬,历经45~50 d;说明在25 ℃下其成花转变阶段提前,且持续时间要短。以上结果均表明,温度能显著影响西红花花芽分化的有无、起始时间和持续长短,同时也能影响其顶芽萌发时间和生长速度,最终影响其开花。
3.2 西红花顶芽中内源激素对花芽分化的调控作用
球根花卉由于其特殊的形态结构,在生理特性、养分运输、环境因子响应等方面均具有一定的特异性,同时具有休眠特性,而休眠的解除一定程度上意味着花芽分化的起始。植物激素是球根休眠和花芽分化的关键诱导因子之一,对花芽分化的起始具有重要的调控作用。IAA对成花的作用是最早被认识到的,但看法不一,有研究认为IAA含量降低有利于花芽分化[23],也有研究认为高含量IAA可能促进花芽分化[24]。目前普遍认为,低浓度IAA是花发生所必需的,但是高浓度则抑制开花。同时,有关ABA和GA3在花芽分化中作用的研究结果[25-26]也不一致,可能因植物不同而有差异。有研究表明,ABA是诱导球根休眠的必需激素[27-28]。郭蕊等[29]对百合的研究结果表明,ABA和GA1+3在解除休眠中起关键作用,其中ABA含量的降低和GA1+3含量的升高可能解除了鳞茎萌发的抑制因素,从而打破休眠、开始花芽分化。郁金香的萌芽生长伴随着IAA和GA含量的增加[30]。GAs对成花的影响可能与处理时期有关,同时对花芽的影响是分阶段的,其含量降低是花芽分化必需的,而在后期花器官发育中,GAs是有促进作用的[31]。
本研究中,IAA含量在花芽分化初期的含量远低于花芽分化前,在随后的形态分化过程中,IAA含量有所增加,但是花芽(25 ℃)IAA含量低于叶芽(15 ℃)IAA含量。该结果与王玉华等[32]对大樱桃的研究结果一致。因此,在西红花顶芽中IAA可能是作为抑花激素。相比于花芽分化前,ABA含量在花芽分化初期降低至谷值,随着花芽分化的进行略有升高,在雄蕊分化期达到峰值后又下降,同时花芽(25 ℃)中ABA含量低于叶芽(15 ℃)。整体说明低水平的IAA和ABA有利于营养生长向生殖生长转变,同时IAA和ABA含量的升高对形态分化有促进作用。同时,花芽(25 ℃)中GA3含量整体呈上升趋势,高于叶芽(15 ℃)中GA3含量,且两者变化趋势相反,说明低水平GA3和适宜的温度下,GA3逐渐积累可能有利于西红花花芽向生殖生长转变,并在后期花器官发育中有促进作用。这与郭蕊等[29]对百合的“ABA含量的降低和GA1+3含量的升高可促进花芽分化”的结果一致。同时,也有文献报道,用适宜浓度GA3浸泡球茎,可以提高西红花花芽分化和开花率,促进开花[33]。
目前,关于激素对花芽形成的影响机理尚不清楚,但普遍认为多种激素之间存在相互促进或相互拮抗的平衡协调效应,其动态的变化和平衡对植物成花起着至关重要的作用。大量研究结果表明,较高的CTKS/GAs、CTKS/IAA、ABA/GAs、ABA/IAA有利于花芽的形成分化和完成[31],但是也有报道称IAA/GAs、ZRs/ABA和IAA/ABA的升高有利于成花[34]。本研究通过比对花芽(25 ℃)和叶芽(15 ℃)中的激素含量比值变化发现,较低的ABA/GA3、IAA/GA3、ABA/IAA有利于西红花顶芽从营养生长向生殖生长转变,且其比值升高可能有利于花芽分化的进行和完成,与花芽分化及成花有密切关系。
本研究中,内源激素变化的结果与张衡锋等[18]对西红花“低水平的GA和IAA有利于花芽分化”的结果一致,但与其“高浓度ABA有利于花芽分化”的结果不一致。可能是由于张衡锋等[18]是根据顶芽解剖学状态的差异来采样,涉及从营养生长期至开花期共6个节点,时间跨度从7月上旬至11月上旬;而本研究是不同温度处理下,选择花芽(25 ℃)和叶芽(15 ℃)两种顶芽发育状态,且主要关注花芽分化转变和花芽分化期,对应西红花顶芽的生长状态有一定的差异。因此,相关研究结果有待进一步验证。
总之,花芽分化是一个复杂的生理生化和形态变化过程,外在环境信号如温度、光照等,内在的营养物质和激素变化等都会对花芽分化起一定的作用。同时,花芽分化并非单一激素变化的结果,而与激素平衡相关,是各种激素相互促进、相互制约的结果。对西红花在自然温度和恒定温度处理下花芽分化进程的观测结果表明,调控温度能有效调控西红花花芽分化的起始和持续长短。花芽分化过程中内源激素含量的变化结果表明,改变外部环境因素也可间接调节内部激素的合成与运输,从而影响休眠与花芽分化。该研究结果可为西红花生产过程中室内环境参数的标准化制定提供理论基础,对其花期调控、生产实践等方面具有一定的指导意义。
猜你喜欢
黑龙江粮食(2022年6期)2022-11-23
中国农学通报(2022年16期)2022-07-08
安徽农业科学(2021年16期)2021-08-30
安徽农业科学(2021年11期)2021-07-16
安徽农学通报(2021年10期)2021-06-17
农民致富之友(2020年22期)2020-09-02
果农之友(2020年5期)2020-06-21
农业与技术(2019年5期)2019-05-29
农民致富之友(2018年21期)2018-12-27
现代农业科技(2016年9期)2016-10-20
