
线粒体活性氧自由基抑制剂R(+)-普拉克索对脑缺血再灌注损伤大鼠JAK2-STAT3通路及炎性因子TNF-α的影响
2021-04-28 01:01黄语悠师文娟赵咏梅刘克建
首都医科大学学报 2021年2期
丁 锚 杨 楠 黄语悠 师文娟 闫 峰 赵咏梅 刘克建
(首都医科大学宣武医院 北京市老年病医疗研究中心 脑血管病转化医学北京市重点实验室,北京 100053)
缺血性脑卒中是导致长期神经功能障碍的重要原因。脑缺血发生时,涉及一系列复杂的病理生理反应,造成内稳态失衡,线粒体功能紊乱。线粒体功能障碍可产生大量活性氧自由基(reactive oxygen species,ROS),进而对神经元造成氧化应激损伤。本课题组研究[1]显示,大鼠脑缺血再灌注6 h时线粒体ROS大量产生,对缺血神经元造成氧化应激损伤,进而引起细胞死亡。这一研究结果首次证明线粒体是脑缺血损伤早期ROS产生的主要来源,线粒体ROS大量生成是造成脑缺血再灌注早期神经元损伤的重要原因之一[1]。多巴胺受体激动剂普拉克索(pramipexole,PPX)具有抗氧化活性,为线粒体靶向抗氧化剂。PPX可通过阻止线粒体通透性转运孔的开放,保护线粒体膜完整性,进而抑制线粒体ROS产生[2]。本团队前期研究[3]表明,R(+)-普拉克索[R(+)-PPX]可显著减少大鼠脑缺血再灌注6 h时线粒体ROS的产生,从而降低大鼠脑梗死体积,说明R(+)-PPX能对脑缺血的早期损伤产生保护作用。但脑缺血再灌注早期R(+)-PPX的神经保护作用机制尚不明确。
内源性酪氨酸激酶2(janus kinase 2,JAK2)-信号转换器和转录激活因子3(signal transducer and activator of transcription 3,STAT3)信号通路是多种细胞因子和生长因子在细胞内传递信号的共同途径,与机体各种生命活动密切相关。JAK2-STAT3通路的激活参与了细胞增殖、炎性反应、血管生成和细胞凋亡等多种生理病理活动[4]。目前关于JAK2-STAT3通路是否在脑缺血再灌注早期发展阶段就被激活的研究尚少。磷酸化STAT3(phosphorylated STAT3,p-STAT3)是STAT3的激活形式,有研究[5]表明,H2O2处理的大鼠成纤维细胞中STAT3被显著激活,并且过表达的p-STAT3可被抗氧化剂抑制,表明JAK2-STAT3信号通路可能在调节氧化应激反应中发挥重要作用。STAT3除了分布在细胞质和细胞核以外,也存在于线粒体中,ROS对心肌细胞线粒体STAT3的氧化作用,使得STAT3被激活[6]。然而,目前暂无研究明确报道,在脑缺血再灌注早期,线粒体ROS是否通过激活JAK2-STAT3通路产生神经损伤作用。
炎性反应是脑缺血再灌注损伤的重要病理生理机制之一,肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)在脑缺血损伤后表达上调,通过促进炎性反应加快细胞凋亡。有文献[7]报道,诱导线粒体ROS产生可增加炎性因子的释放,提示线粒体ROS可直接调节炎性因子的表达。另外,JAK2-STAT3也是一条重要的炎性反应相关通路,JAK2-STAT3通路的激活参与炎性反应相关的信号转导过程。研究[8]显示,应用JAK2抑制剂AG490可抑制STAT3活化,进而明显下调成纤维细胞内TNF-α的表达水平,说明JAK2-STAT3通路的激活也能影响TNF-α的表达。
为了深入探究R(+)-PPX保护脑缺血再灌注早期损伤的机制,本研究采用大脑中动脉梗塞(middle cerebral artery occlusion,MCAO)再灌注模型大鼠,观察缺血再灌注6 h时缺血脑组织p-JAK2、p-STAT3和TNF-α的表达变化,以及R(+)-PPX对缺血大鼠脑内JAK2-STAT3通路和TNF-α表达的影响,以期为R(+)-PPX的脑保护作用研究提供更多的科学依据。
1 材料与方法
1.1 实验动物及分组
健康雄性SD大鼠体质量为280~320 g,购自北京维通利华实验动物公司,实验动物许可证号:SCXK(京)2012-0001,于SPF级动物实验室恒温饲养,术前12 h禁食水。将30只大鼠应用数字表法随机分为3组:假手术组(Sham组)、MCAO再灌注6 h组(MCAO组)和MCAO再灌注6 h+R(+)-PPX组[R(+)-PPX组]。每组10只大鼠。R(+)-PPX溶于0.9%(质量分数)氯化钠注射液(以下简称生理盐水)中。R(+)-PPX组于MCAO前30 min腹腔一次性注射R(+)-PPX,剂量为1 mg/kg[1],Sham组和MCAO组皮下注射同等体积的生理盐水。
1.2 主要仪器设备
小动物麻醉机(Harvard Apparatus公司,美国)、手术显微镜(Carl Zeiss公司,德国)、反馈式温度调节仪(CMA 150,Carnegie Medicin公司,瑞典)、双极电凝器(ACC100,北京德威医疗器械有限公司)、小动物呼吸机(Harvard Apparatus 683,Harvard Apparatus公司,美国)、冰冻切片机(Thermo Fisher Scientific公司,美国)、荧光显微镜(Nikon公司,日本)。
1.3 试剂
恩氟烷(河北一品制药有限公司);水合氯醛(首都医科大学宣武医院); R(+)-PPX(Sigma公司,美国);DHE(Sigma公司,美国);p-JAK2抗体(CST公司,美国); p-STAT3抗体(CST公司,美国);TNF-α抗体(CST公司,美国);β-actin抗体(Santa Cruz公司,美国)等。
1.4 动物模型制作
MCAO大鼠模型的制备依照改良Zea Longa法。在70%(体积分数)N2O和30%(体积分数)O2中混合5%(体积分数)恩氟烷诱导麻醉,而后改用面罩吸入2%(体积分数)恩氟烷维持麻醉。大鼠仰卧位固定,去除颈部毛并消毒后,沿颈部正中线做一纵行切口,分离且夹闭右侧颈总动脉。分离右侧颈外动脉,结扎并电凝断后,眼科剪呈 45°在右侧颈外动脉近心端血管壁上剪一小口,将尼龙线栓(头端直径为0.38 mm)插入至大脑中动脉起始端处固定线栓。术中使用反馈性控温毯监测术中大鼠肛温,使其维持在(37.0±0.5)℃,并检测大鼠心率及血压。于缺血90 min 后拔出线栓,进行再灌注。Sham组大鼠相应部位皮肤切开分离血管后不插栓即缝合。
1.5 DHE染色
Sham组、MCAO组以及R(+)-PPX组大鼠腹腔注射水合氯醛,快速断头取脑包埋,行脑组织冰冻切片,每片脑组织厚度均为20 μm。未进行固定的新鲜冰冻切片浸入10 μmol/L的超氧化物阴离子荧光探针(dihydroethidium,DHE)溶液中(生理盐水溶解),避光条件下37 ℃孵育90 min后用PBS冲洗,封片后于荧光显微镜下观察,观察区域为缺血侧脑皮质中的半暗带区。
1.6 免疫荧光双标染色
各组大鼠脑组织冰冻切片(厚度20 μm)于预冷的4%(质量分数)多聚甲醛中固定10 min,PBS漂洗后用含0.2%(体积分数)TritonX-100的PBS室温孵育10 min,之后滴加5%(体积分数)山羊血清室温封闭30 min,弃血清,滴加一抗(p-STAT3抗体 1∶100,TNF-α抗体 1∶100),4 ℃孵育过夜。次日将切片于室温复温1 h,PBS漂洗3次,滴加荧光二抗(1∶200稀释),室温避光孵育1 h。PBS漂洗3次,滴加含DAPI(4′,6-diamidino-2-phenylindole)的封片剂,之后在荧光显微镜下观察。
ROS分别与p-STAT3、TNF-α的荧光双标染色:脑组织冰冻切片用DHE荧光染剂孵育90 min。PBS冲洗后,滴加一抗(p-STAT3抗体1∶100,TNF-α抗体 1∶100)孵育过夜。次日将切片室温复温1 h,PBS漂洗后,滴加荧光二抗(1∶200稀释)。用含DAPI的封片剂封片后在荧光显微镜下观察。
1.7 Western blotting检测
取新鲜大鼠右侧脑组织,立即于前囟0~-0.1处切取1 mm厚的脑组织冠状切片。加入含有蛋白酶抑制剂的裂解液后,将缺血侧脑组织于冰上匀浆,冰浴静置30 min后,4 ℃条件下12 000 g离心30 min取上清。用BCA法测定蛋白浓度。样品中加入上样缓冲液煮沸,采用SDS-PAGE电泳湿转法转膜90 min,于5%(质量分数)脱脂牛奶室温下封闭1 h,加入一抗(p-JAK-2抗体1∶1 000,β-actin抗体1∶1 000)孵育过夜,次日用TBST洗膜3次,室温下于HRP标记的山羊抗小鼠/兔(1∶6 000)二抗中孵育1 h,TBST洗膜3次,用ECL化学发光液显色,并用化学发光系统扫描检测。
1.8 统计学方法

2 结果
2.1 R(+)-PPX抑制MCAO大鼠缺血侧脑组织p-JAK2的表达
Western blotting检测结果显示,3组间大鼠右侧脑组织p-JAK2表达量差异有统计学意义(P<0.05)。两两比较结果显示,与Sham组相比,MCAO组大鼠再灌注6 h缺血侧脑组织p-JAK2表达量明显增加;与MCAO组大鼠相比,R(+)-PPX组大鼠再灌注6 h缺血侧脑组织p-JAK2表达量显著降低,差异有统计学意义(P<0.05)(图1)。

图1 蛋白免疫印迹法检测Sham、MCAO及R(+)-PPX组大鼠再灌注6 h缺血脑组织p-JAK2蛋白的表达Fig.1 Expression of p-JAK2 in ischemic brain tissue of Sham, MCAO and R(+)-PPX groups after 6 h reperfusion Representative Western blotting images of p-JAK2 and quantification of optical density measurements normalized to β-actin. Data are (n=6). *P<0.05 vs Sham group, #P<0.05 vs MCAO group; MCAO: middle cerebral artery occlusion; PPX: pramipexole; p-JAK2: phosphor-ylated-janus kinase 2.
2.2 R(+)-PPX抑制MCAO大鼠脑缺血半暗带区p-STAT3的表达
免疫荧光染色结果显示,Sham组大鼠与缺血半暗带相对应的脑区未见p-STAT3绿色荧光阳性细胞,而MCAO、R(+)-PPX组大鼠再灌注6 h缺血半暗带区均可见p-STAT3阳性细胞(图2A),3组间大鼠缺血半暗带区p-STAT3阳性细胞数差异有统计学意义(P<0.05)。两两比较结果显示,与Sham组相比,MCAO组大鼠再灌注6 h缺血半暗带区p-STAT3阳性细胞数明显增加;与MCAO组相比,R(+)-PPX组大鼠再灌注6 h缺血半暗带区p-STAT3阳性细胞数明显减少,差异有统计学意义(P<0.05)(图2B)。
2.3 脑缺血再灌注早期,大鼠脑缺血半暗带区ROS与p-STAT3共定位
荧光双标结果显示,MCAO组大鼠再灌注6 h缺血半暗带区ROS荧光探针DHE呈现红色荧光,p-STAT3免疫荧光染色阳性细胞呈现绿色荧光,DAPI染色细胞核为蓝色荧光。进行图像合并后发现,DHE的红色荧光阳性细胞和p-STAT3的绿色荧光阳性细胞重合,说明ROS与p-STAT3共定位(图2A)。提示脑缺血再灌注早期,ROS的产生与STAT3的激活相关。
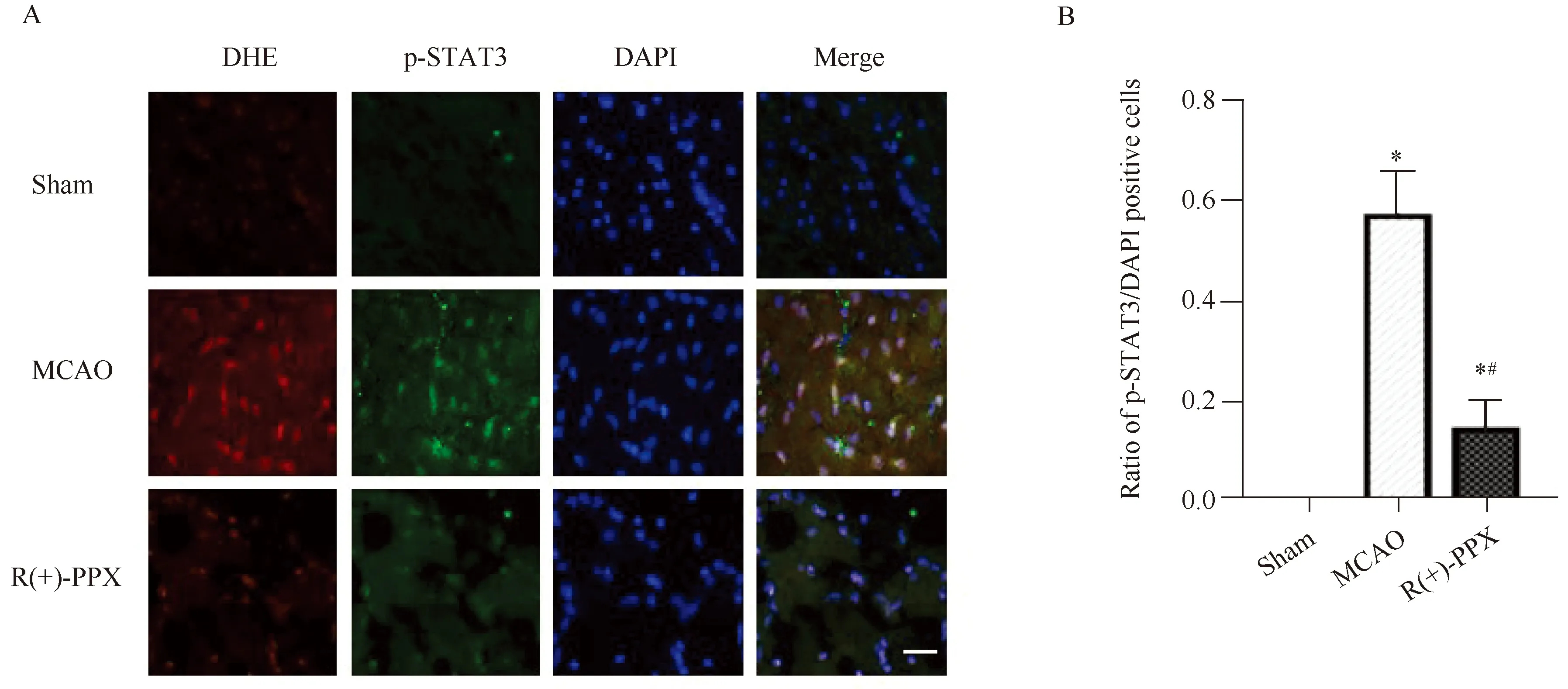
图2 荧光双标染色法检测Sham、MCAO与R(+)-PPX组大鼠再灌注6 h脑缺血半暗带区p-STAT3和ROS的表达Fig.2 Expression of p-STAT3 and ROS in the ischemic penumbra of Sham, MCAO and R(+)-PPX groups after 6 h reperfusionA: representative double labeling fluorescence images for the colocalization of ROS (red) and p-STAT3 (green) after 6 h reperfusion. Scale bar=20 μm; B: quantification of p-STAT3 positive cells. Data are (n=6). *P<0.05 vs Sham group, #P<0.05 vs MCAO group; MCAO: middle cerebral artery occlusion; PPX: pramipexole; p-STAT3: phosphorylated signal transducer and activator of transcription 3; ROS: reactive oxygen species; DAPI: 4′,6-diamidino-2-phenylindole.
2.4 脑缺血再灌注早期,大鼠脑缺血半暗带区ROS与炎性因子TNF-α共定位,且R(+)-PPX减少MCAO大鼠脑缺血半暗带区TNF-α的表达
荧光染色结果显示,MCAO组大鼠再灌注6 h缺血半暗带区ROS荧光探针DHE呈现红色荧光,TNF-α免疫荧光染色阳性细胞呈现绿色荧光,DAPI染色细胞核为蓝色荧光。进行图像合并后发现,DHE的红色荧光阳性细胞和TNF-α的绿色荧光阳性细胞重合,说明ROS与TNF-α共定位(图3A)。
Sham组大鼠与缺血半暗带相对应的脑区未见TNF-α绿色荧光阳性细胞,而MCAO、R(+)-PPX组大鼠再灌注6 h缺血半暗带区均可见TNF-α阳性细胞(图3A),3组间大鼠缺血半暗带区TNF-α阳性细胞差异有统计学意义(P<0.05)。两两比较结果显示,与Sham组相比,MCAO组大鼠再灌注6 h缺血半暗带区TNF-α阳性细胞数明显增加;与MCAO组大鼠相比,R(+)-PPX组大鼠再灌注6 h缺血半暗带区TNF-α阳性细胞数明显减少,差异有统计学意义(P<0.05)(图3B)。
2.5 脑缺血再灌注早期,大鼠脑缺血半暗带区p-STAT3与TNF-α共定位
免疫荧光双标染色结果显示,MCAO组大鼠再灌注6 h缺血半暗带区p-STAT3呈现红色荧光,TNF-α免疫荧光染色阳性细胞呈现绿色荧光,DAPI染色细胞核为蓝色荧光。进行图像合并后发现,p-STAT3阳性细胞的红色荧光和TNF-α阳性细胞的绿色荧光重合,说明脑缺血再灌注早期,大鼠脑缺血半暗带区p-STAT3与TNF-α共定位(图3C)。

图3 荧光双标染色法检测Sham、MCAO与R(+)-PPX组大鼠再灌注6 h脑缺血半暗带区TNF-α和ROS的表达;免疫荧光双标染色法检测MCAO大鼠脑缺血半暗带区p-STAT3和TNF-α的表达Fig.3 Expression of TNF-α and ROS in the ischemic penumbra of Sham, MCAO and R(+)-PPX groups after 6 h reperfusion;expression of p-STAT3 and TNF-α in the ischemic penumbra of MCAO ratsA: representative double labeling fluorescence images for the colocalization of ROS (red) and TNF-α (green) after 6 h reperfusion. Scale bar=20 μm; B: quantification of TNF-α positive cells. Data are (n=6). *P<0.05 vs Sham group, #P<0.05 vs MCAO group; C: representative double labeling immunofluorescence images of p-STAT3 (red) and TNF-α (green) after 6 h reperfusion. Scale bar=20 μm; MCAO: middle cerebral artery occlusion; PPX: pramipexole; TNF-α: tumor necrosis factor-α; ROS: reactive oxygen species; p-STAT3: phosphorylated STAT3; DAPI: 4′,6-diamidino-2-phenylindole.
3 讨论
氧化应激是脑缺血再灌注过程中加重脑组织损伤的重要原因之一。在脑缺血后,特别是再灌注发生时,神经元内产生大量自由基、过氧化物等造成细胞氧化应激损伤[9]。线粒体是体内超氧化物的主要来源之一,本课题组最近的研究[1]显示,脑缺血再灌注早期的氧化应激损伤主要是由于线粒体ROS大量产生造成的,因此抑制线粒体ROS可能对早期脑缺血再灌注损伤具有神经保护作用。PPX是线粒体靶向抗氧化剂,能抑制线粒体ROS的产生。笔者在实验中证明,R(+)-PPX可明显减少脑缺血再灌注6 h大鼠缺血大脑半球线粒体ROS的生成,从而发挥神经保护作用[3]。然而,R(+)-PPX在缺血性脑损伤过程中发挥早期脑保护作用的具体机制尚不完全清楚。
JAK2-STAT3作为重要的细胞因子信号转导途径,参与细胞增殖分化、氧化应激、炎性反应、免疫调节等多个生理病理环节。有研究[10]显示,JAK2-STAT3通路在脑缺血再灌注24 h后被激活,说明JAK2-STAT3通路参与脑缺血损伤。但目前关于JAK2、STAT3是否在脑缺血再灌注早期即被激活的研究尚少。笔者利用Western blotting检测脑缺血大鼠再灌注6 h后缺血侧脑组织中p-JAK2表达量,免疫荧光染色法检测p-STAT3的水平,研究结果显示,MCAO组大鼠再灌注6 h,缺血半暗带区p-JAK2及p-STAT3表达量比Sham组明显增加,说明 JAK2-STAT3通路在脑缺血再灌注早期即被激活。
研究[5,11]证明,JAK2-STAT3信号通路在氧化应激损伤的细胞及动物模型中被激活,提示该信号通路可能在调节氧化应激反应中发挥重要作用。有文献[12]报道,ROS作为一种信号分子可激活JAK2,进而导致STAT3磷酸化。而清除活性氧能抑制体内STAT3磷酸化。由此可见,ROS对于启动JAK2-STAT3通路具有重要意义。为了探究ROS在脑缺血再灌注早期能否介导JAK2-STAT3通路的激活。笔者对MCAO再灌注6 h大鼠缺血半暗带区ROS与p-STAT3进行荧光双标染色,结果显示ROS与p-STAT3共定位,说明脑缺血再灌注6 h时,STAT3的激活与ROS的产生密切相关。笔者还发现,腹腔注射R(+)-PPX抑制ROS后,大鼠缺血脑组织内p-JAK2、p-STAT3表达水平均显著下降。该结果进一步说明,脑缺血再灌注早期,线粒体ROS的大量产生可激活JAK2-STAT3通路。结合本课题组之前的研究[3]结果,给予R(+)-PPX治疗能明显降低MCAO再灌注6 h大鼠脑梗死体积,减少细胞凋亡,笔者推测,R(+)-PPX很可能通过抑制JAK2-STAT3通路的激活,从而发挥神经保护作用。
炎性反应是加重脑缺血再灌注损伤的重要原因之一,脑缺血急性期大量释放的TNF-α是缺血后炎性损伤的重要启动因子[13]。有研究[14]显示,线粒体ROS参与铜诱导小胶质细胞炎性反应的过程,抑制ROS可缓解铜导致的炎性反应,降低炎性因子TNF-α的水平。为了观察脑缺血再灌注损伤后TNF-α的表达是否与ROS有关,本研究对TNF-α和ROS进行荧光染色双标,结果显示,大鼠缺血半暗带区TNF-α阳性细胞与ROS共定位,说明脑缺血再灌注早期TNF-α的表达与ROS密切相关。给予R(+)-PPX抑制线粒体ROS产生后可明显降低MCAO再灌注6 h大鼠脑缺血半暗带区TNF-α的表达量,进一步说明,脑缺血再灌注早期线粒体ROS能直接诱导炎性因子TNF-α的表达。另外,研究[10]报道,JAK2-STAT3通路被激活后,p-STAT3与细胞核DNA结合从而增加炎性因子的表达。对周围巨噬细胞的研究[15]也发现,JAK2-STAT3的激活能介导炎性因子的释放,诱发巨噬细胞炎性反应。本研究用免疫荧光染色法对p-STAT3和TNF-α进行双标,发现p-STAT3阳性细胞和TNF-α阳性细胞在大鼠脑缺血半暗带区能够重合,说明脑缺血再灌注损伤后TNF-α的产生与p-STAT3的激活有关。因此,笔者推测,脑缺血再灌注损伤后线粒体ROS也可能是通过激活JAK2-STAT3通路,进而引起炎性因子TNF-α的表达增加。
了解线粒体ROS损伤神经元的机制,对于早期挽救缺血半暗带具有重要意义。本研究重点探究了脑缺血再灌注损伤早期,R(+)-PPX发挥抗氧化作用的机制。证明R(+)-PPX通过减少MCAO再灌注6 h大鼠线粒体ROS产生,抑制JAK2-STAT3通路激活,降低TNF-α表达,产生神经保护作用。该结果在本课题组前期研究的基础上,进一步完善了线粒体ROS造成脑缺血再灌注早期损伤的机制,并为深入探讨R(+)-PPX的神经保护作用机制提供了相关的实验数据。
猜你喜欢
山西医科大学学报(2022年9期)2022-11-08
中国现代医生(2022年23期)2022-09-21
中华胰腺病杂志(2022年4期)2022-08-23
中国中医药信息杂志(2022年6期)2022-06-27
健康体检与管理(2022年4期)2022-05-13
昆明医科大学学报(2022年3期)2022-04-19
中国医药导报(2017年11期)2017-06-01
医学信息(2016年35期)2017-02-23
中国民族民间医药·下半月(2015年12期)2016-02-25
江苏农业科学(2014年8期)2014-10-23
