
MaFGF对慢性脑缺血再灌注损伤小鼠神经功能保护的机制研究
2021-04-25 10:21李燕陈克龙张妤婷支海鸯黎斌孙慧静
浙江临床医学 2021年3期
李燕 陈克龙 张妤婷 支海鸯 黎斌 孙慧静 陈 凌*
缺血性脑卒中是最常见的一种具有较高的致残率、致死率和复发率的脑血管疾病,且发病率逐年上升,呈年轻化趋势[1]。及时恢复缺血区的血供以保护缺血半暗带神经元是治疗关键,但脑缺血后血管再通,可导致缺血再灌注损伤,引起缺血及其周围神经细胞坏死和凋亡,加重脑缺血损伤。改构型酸性成纤维细胞生长因子(MaFGF)是一种多功效的生长刺激因子,具有调节神经、舒张血管、保护神经等多种生物学效应[2]。目前研究已证实MaFGF具有良好的抗脑缺血再灌注损伤作用[3],但其对慢性脑缺血损伤神经保护的作用机制尚不明确。本研究通过观察MaFGF对慢性脑缺血再灌注损伤小鼠PI3K/AKT信号通路及内质网应激凋亡通路相关蛋白的影响,探讨MaFGF的神经保护功能及其作用机制。
1 材料与方法
1.1 实验动物及材料 2~3月龄雄性C57BL/6J小鼠,体重22~25 g,购自上海斯莱克动物实验中心,动物许可证号为SCXK(沪)2012-0002。MaFGF(暨南大学生物工程研究所),PI3K抗体(abcam,ab86714)、p-PI3K抗体(abcam,ab182651)、p-AKT抗体(abcam,ab38449)、AKT抗体(abcam,ab8805)、caspase-12抗体(abcam,ab62484)、CHOP抗体(abcam,ab11419)、GRP170抗体(abcam,ab241049),山羊抗小鼠IgG/TRITC(中杉金桥公司,货号ZF-0313)红光二抗、山羊抗小鼠IgG/FITC(中杉金桥公司,ZF-0312)绿光二抗,Morris水迷宫(上海吉量软件科技公司),化学发光凝胶成像系统(北京东胜创新生物科技公司),荧光显微镜成像系统(OLYMPUS公司,DP72)。
1.2 建模与分组 24只雄性C57BL/6J小鼠饲养于SPF级动物房(室温22±2℃,湿度50±5%)。按照随机数字法将实验小鼠分为Sham组、BCAO组和BCAO+MaFGF组,每组各8只。双侧颈动脉夹闭获得脑缺血再灌注损伤模型:麻醉小鼠(10%水合氯醛),以仰卧姿势固定于手术台上,置于加热毯上保持直肠温度为37℃左右,同时保持呼吸顺畅,在颈部腹侧中线处常规消毒后做1 cm左右的切口,钝性分离双侧颈总动脉,夹闭双侧颈总动脉约30 min,去除动脉夹恢复灌注,缝合切口;Sham组仅切开皮肤;再灌注后,BCAO+MaFGF组开始鼻内给药20 μg/kg MaFGF,其余2组给予等体积的生理盐水,1次/d,连续28 d。
1.3 神经功能评估 分别于术前/术后第28天,采用神经功能缺损评分(mNSS)评估所有实验小鼠的神经功能损害程度,包括运动、感觉、反射、平衡试验四个方面,分值为0~18分,小鼠无神经功能损伤体征则评为0分。
1.4 水迷宫测试 (1)定位航行试验:所有小鼠在给药结束后开始训练,将平台固定于目标象限的中央,圆池中注入清水(高出平台约2~3 cm),在4个象限各取一点作为入水口并依次放入小鼠。记录60 s内小鼠的运动轨迹及逃避潜伏期(从入水到爬上平台的时间,上限为60 s)。每天在同一时间训练小鼠,连续训练5 d。(2)空间探索试验:在训练第6天,移除平台,从4个相同入水点依次放入小鼠,观察并记录60 s内小鼠的运动轨迹及穿越原平台位置次数,分析计算平均速度及在目标象限停留的时间百分比。
1.5 样本采集与保存 取0.9%氯化钠溶液心脏灌注排出脑组织血液,快速断头,取出全脑组织,放入液氮中速冻,用于后续蛋白测定及制备冰冻切片。部分小鼠继以4%多聚甲醛心脏灌注直至小鼠全身僵硬,取出大脑浸入4%多聚甲醛液中固定,进行石蜡包埋,切片,低温保存备用。
1.6 Nissl染色 取石蜡切片(片厚4 μm)进行常规脱蜡、水化、洗涤,加入甲苯胺蓝溶液染色(60 ℃,40 min),随后进行脱水(梯度乙醇)、透明(二甲苯溶液)、封片(透明树脂),光学显微镜下观察各种小鼠海马CA1区神经元形态学变化。
1.7 免疫荧光染色 液氮速冻脑组织,切片机切片,片厚8 μm,35 ℃烘箱复温5 min,4%多聚甲醛固定10 min,封闭液孵育30 min;将p-AKT、GRP170一抗稀释液(1:100)混合后覆盖各标本,置于湿盒内4 ℃孵育过夜,PBS漂洗3次;荧光二抗室温下避光孵育1 h,PBS洗涤;最后DAPI孵育5 min,PBS洗涤,抗荧光猝灭剂封片;荧光显微镜下,随机选取5个视野,观察各组小鼠大脑海马CA1区神经元p-AKT及GRP170的表达及位置情况,并拍照。所有步骤均在黑暗中进行。
1.8 Western blot RIPA组织裂解液裂解海马组织细胞,提取总蛋白,使用BCA试剂盒测定各组蛋白浓度,制备蛋白样品。使用10%SDS-聚丙烯酰胺凝胶电泳,将各组蛋白样品中不同分子大小的蛋白分离并转移至聚偏二氟乙烯(PVDF)膜上,转膜完成后脱脂奶粉室温封闭1 h。随后一抗稀释液4 ℃孵育过夜,滴加二抗稀释液室温孵育2 h,最后条带滴加ECL化学曝光液,通过Bio-rad曝光机显色曝光,采用ImageJ分析各蛋白灰度值,比较各组小鼠大脑海马区p-PI3K、p-AKT、caspase-12、CHOP、GRP170的蛋白表达水平。
1.9 统计学方法 采用SPSS 18.0统计软件。计量资料以(±s)表示,多组之间比较采用单因素方差分析,两两比较采用LSD法检验。以P<0.05为差异有统计学意义。
2 结果
2.1 各组小鼠的mNSS评分比较 各组小鼠术前mNSS评分均为0分,表明术前各组小鼠的神经功能均正常;术后Sham组小鼠评分为0分,BCAO组小鼠mNSS评分与Sham组比较明显升高,差异有统计学意义(P<0.05);给予MaFGF治疗后,BCAO+MaFGF组小鼠mNSS评分明显降低,差异有统计学意义(P<0.05)。见图1。
2.2 各组小鼠认知行为能力比较 在定位航行试验中,随着训练次数的增加,各组小鼠的逃避潜伏期逐渐降低。在训练第3天,与Sham组比较,BCAO组小鼠逃避潜伏期明显下降(P<0.05);与BCAO组比较,MaFGF组小鼠逃避潜伏期均明显缩短(P<0.05)。在空间探索试验中,BCAO组小鼠平均穿越平台次数与Sham组比较明显减少(P<0.05);MaFGF组平均穿越平台次数及60 s内在原平台象限停留的时间百分比与BCAO组比较明显增加(P<0.05)。见图2。
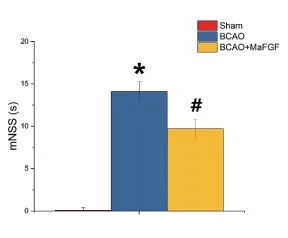
图1 术后各组小鼠mNSS评分结果。与Sham组比较,★P<0.05;与BCAO组比较,# P<0.05

图2 A 各组小鼠在定位航行试验中训练第5天的运动轨迹;B 各组小鼠在空间探索实验中穿越平台的运动轨迹;C 各组小鼠逃避潜伏期变化趋势及空间探索实验结果,包含平均速度(cm/s)、穿越平台次数(times)和目标象限时间(s);与Sham组比较,★ P<0.05;与BCAO组比较,# P<0.05
2.3 各组小鼠海马CA1区神经元细胞形态结构的变化 Nissl染色显示,Sham组小鼠海马CA1区神经元细胞呈圆形或锥体形,细胞排列整齐,细胞核饱满,核仁明显,胞浆呈深蓝色,内含丰富的尼氏体;与Sham组比较,BCAO组小鼠海马CA1区神经元细胞排列紊乱、疏松,可见大量细胞结构破坏,细胞核固缩、碎裂,坏死细胞数目明显增多(P<0.05);与BCAO组比较,BCAO+MaFGF组小鼠海马CA1区正常神经元细胞增多,坏死细胞数目明显减少(P<0.05)。见图3。

图3 各组小鼠海马CA1区神经元细胞Nissl染色(×200);与Sham组比较,★ P<0.05;与BCAO组比较,# P<0.05
2.4 各组小鼠海马区p-PI3K、p-AKT、GRP170、CHOP、caspase-12蛋白表达水平比较 BCAO组小鼠海马内CHOP和caspase-12与Sham组相比明显升高(P<0.05),p-PI3K、p-AKT、GRP170均未见明显改变(P>0.05);经MaFGF处理后,CHOP和Caspase-12明显降低,而p-PI3K、p-AKT、GRP170表达明显增加(P<0.05)。见图4。

图4 各组小鼠海马CA1区p-PI3K、p-AKT、GRP170、Caspase-12、CHOP蛋白表达水平比较;与Sham组比较,★ P<0.05;与BCAO组比较,# P<0.05
2.5 各组小鼠海马区p-AKT、GRP170免疫阳性颗粒的表达情况 p-AKT和GRP170免疫阳性颗粒均聚集在海马神经元胞浆内;BCAO组小鼠海马区p-AKT和GRP170免疫阳性颗粒表达量与Sham组比较差异无统计学意义;GRP170和p-AKT在BCAO+MaFGF组的分布均比BCAO组更趋致密。见图5。
3 讨论
酸性成纤维细胞生长因子(aFGF,FGF-1)是一种多功能生长刺激因子,因aFGF的特殊结构赋予其在血管再生、创伤修复、骨骼生长、神经保护等方面显示出卓越的临床应用价值,也成为了基础研究领域的热点[2]。但由于aFGF有丝分裂活性可能与细胞增殖、分化和肿瘤发生有一定的关系,使aFGF的应用受到一定限制。因此,本研究采用改构型aFGF观察其对慢性脑缺血再灌注损伤小鼠的神经保护作用。aFGF的N端第21~27位的氨基酸Asn-Tyr-Lys-lys-Pro-Lys-Leu是aFGF促有丝分裂活性的重要氨基酸序列,定向敲除该段序列的第23~26位氨基酸片段,由Met取代Leu,可使其失去促有丝分裂能力,同时保留了非促分裂活性结构域[4]。XU等[3]研究表明MaFGF明显减轻局灶性缺血再灌注大鼠的脑水肿程度和脑梗死面积,降低丙二醛含量,升高超氧化物歧化酶活性,对急性局灶性缺血再灌注大鼠发挥神经保护作用。本研究结果显示,MaFGF可明显降低BCAO组小鼠28 d时的mNSS评分,缩短逃避潜伏期,增加穿台次数,表明MaFGF能够改善慢性脑缺血再灌注损伤小鼠的神经功能和认知记忆力,发挥长期的神经保护作用。

图5 各组小鼠海马CA1区p-AKT、GRP170免疫阳性颗粒表达情况(×200)
脑缺血再灌注后,神经元细胞损伤主要表现为细胞凋亡,因此细胞凋亡是脑缺血再灌注损伤的中心环节和治疗主要靶点。内质网应激(ERS)、钙超载、氧化应激、炎性反应等诸多因素,均可导致细胞凋亡,其中内质网应激介导的细胞凋亡具有关键作用。当ERS过强或持续存在时,一方面内质网功能无法恢复,使细胞失衡转向凋亡,一方面诱导激活增强剂结合蛋白同源蛋白(CHOP)、c-Jun 氨基末端激酶(JNK)和胱氨酸天冬氨酸特异性蛋白酶12(Caspase12)等激活细胞凋亡通路,通过凋亡清除受损严重的细胞。本研究结果显示,与Sham组比较,BCAO小鼠海马CA1区CHOP、Caspase-12表达明显升高,神经元细胞形态结构破坏严重,提示脑缺血再灌注损伤小鼠海马神经元细胞存在内质网功能障碍。葡萄糖调节蛋白170(GRP170),是驻留在ER中的主要分子伴侣,与GRP78具有相似的调控作用。有文献报道GRP170可以作为一种核苷酸交换因子(NEF),触发结合免疫球蛋白(BiP)核苷酸转录形成ATP-BiP,除了其公认的NEF活性外,其还包含一个C端保持酶阈,能够促进未折叠或错误折叠蛋白的降解,减轻内质网应激,从而起保护作用[5-6]。因此GRP170的高表达可以促进内质网功能的稳定,减少内质网应激相关细胞凋亡。本研究结果显示,与Sham组比较,BCAO小鼠海马区GRP170表达无明显改变,这可能是在发生ERS初期,GRP170应激性增多以激活未折叠蛋白反应(UPR),恢复内质网的功能;但ERS持续存在,使得GRP170大量消耗而逐渐减少。aFGF可通过抑制内质网应激发挥神经保护作用,HU等[7]应用aFGF鼻内给药1周持续治疗新生儿缺血缺氧型脑损伤,可显著减少脑梗死面积,通过抑制内质网应激发挥长期的神经保护作用。本研究结果显示MaFGF可上调BCAO小鼠海马区神经元细胞GRP170的表达,恢复内质网功能,降低CHOP、Caspase-12的表达水平,减少神经元凋亡。
FGF可通过与FGF受体结合,促使FGF受体磷酸化并产生酪氨酸蛋白激酶活性,启动PI3K途径、蛋白激酶C(PKC)途径、丝裂原活化蛋白酶(MAPK)途径等发挥其生物学效应,其中PI3K/AKT通路是aFGF在神经组织中发挥作用的主要信号途径[8-9]。外源性aFGF可通过激活PI3K-Akt-Rac1信号通路部分降低小分子G蛋白RhoA活性,从而改善脑外伤后的神经功能缺陷和保持血脑屏障完整性[10]。WANG等[11]研究发现bFGF可作用于靶细胞诱发PI3K信号通路级联反应,激活下游ERK1/2、AKT、GSK3β通路,抑制内质网应激反应(ERS),从而减少神经组织病变和神经细胞凋亡。本研究显示,MaFGF可增加BCAO小鼠神经元细胞PI3K、AKT的磷酸化,表明MaFGF可能是通过激活PI3K/AKT信号通路级联反应而发挥抗凋亡作用。
综上,MaFGF能够明显改善慢性脑缺血再灌注损伤小鼠的神经功能,改善认知和记忆能力,其可能机制是通过激活PI3K/AKT信号通路,减轻内质网应激,减少神经元细胞凋亡从而发挥神经保护作用。
猜你喜欢
医学研究生学报(2022年3期)2022-11-27
健康体检与管理(2022年4期)2022-05-13
作文周刊·小学二年级版(2022年20期)2022-05-05
昆明医科大学学报(2022年3期)2022-04-19
医学研究杂志(2021年12期)2021-11-30
现代临床医学(2021年1期)2021-01-26
智慧健康(2020年20期)2020-12-05
医学信息(2016年35期)2017-02-23
科技知识动漫(2016年6期)2016-06-24
汽车观察(2009年1期)2009-02-18
