
光强对4种鸭跖草科植物叶片生理特性和超微结构的影响
2021-04-08 10:55李子葳杨宇佳杨小梅
草地学报 2021年2期
陈 斌, 薛 晴, 李子葳, 杨宇佳, 杨小梅, 薄 杉, 李 强, 何 淼
(东北林业大学园林学院, 黑龙江 哈尔滨 150040)

紫鸭跖草(Commelinapurpurea)、‘花叶’水竹草(Tradescantiaflurnuensis‘Variegata’)、吊竹梅(Tradescantiazebrina)、‘绿叶’水竹草(Tradescantiaflurnuensis‘Vairidia’)是鸭跖草科(Commelinaceae)的4种植物,植株低矮、叶色丰富、耐瘠薄、茎多呈匍匐状、茎节处易生气生根、繁殖能力强、成坪覆盖速度快且价格低廉,现已经作为地被植物在园林绿化中应用,具有较强的光适应能力,但每种植物的适光策略目前尚不清晰。因此,本试验通过人工模拟不同立地条件下的光强,通过遮光网设置5种光环境对紫鸭跖草、‘花叶’水竹草、吊竹梅和‘绿叶’水竹草进行光处理,测定不同光强下4种植物的生理生化指标和叶肉细胞的超微结构,揭示4种植物的适光策略和潜能,以期为其在园林中栽培应用和进一步大范围的推广提供科学依据。
1 材料和方法
1.1 试验材料
试验于2019年3—7月在黑龙江省哈尔滨市东北林业大学花卉研究所苗圃进行。以紫鸭跖草、‘花叶’水竹草、吊竹梅、‘绿叶’水竹草为材料(表1),3月初选取当年生健壮的枝条,剪取顶部长约5 cm的茎段作为插条,基质采用75%腐殖土+25%蛭石混合,经120℃高温灭菌后装入直径12 cm高18 cm的花盆中,采用浸盆法充分浸润盆中的栽培基质,然后进行扦插。每盆扦插2个茎段,每种试验材料各60盆。扦插完毕后放置在育苗室内缓苗,缓苗期间的环境条件为:温度24~26℃,相对湿度60%~70%,12 h光照,白天光强为480 μmol·m-2·s-1。缓苗1个月后,选取长势旺盛,无病虫害的植株进行试验。

表1 植物材料
1.2 试验方法
于2019年4月30在苗圃内利用不同密度的黑色尼龙遮光网设置5种光强梯度(分别为自然光强的100%,75%,50%,25%,5%),每个处理间隔50 cm,顶部遮光网距植株体顶端30 cm,每个光强梯度每种植物材料各处理12盆,3个重复。每隔5 d浇水一次,处理45 d后,进行各项指标的测定。
1.3 指标测定

二氨基联苯胺法(diaminobenzidine,DAB)染色测定H2O2:摘取茎顶部第3~5片成熟完整的叶片,放入pH为5.8的DAB染色液(1 mg·mL-1)中,在28℃下过夜避光保存,然后放入80%的乙醇中水浴煮沸若干次,直至叶片颜色完全脱去,并用相机拍照。
叶片超微结构的观察:取健壮植株的第3片功能叶,在叶脉两侧各取1 mm×3 mm的矩形小块,每组处理取3片。用浓度为2.5%的戊二醛溶液和1%的锇酸进行双固定,固定后用pH6.8的磷酸缓冲液漂洗;然后用不同浓度的乙醇及丙酮溶液依次脱水,并用环氧树脂包埋;采用LKB-5型超薄切片机进行切片,切片经醋酸铀-枸橼酸铅双染色后,于H7650型透射电子显微镜下对材料进行观察并拍照[17],每个处理取10个视野,统计每个视野中每个叶肉细胞内的叶绿体、线粒体、淀粉粒数量,并在每个视野中随机选取5个叶绿体,统计每个叶绿体中嗜锇颗粒数量。
1.4 数据整理与分析
用Excel 2016对数据进行整理和作图,利用SPSS 22.0对数据进行方差分析,用Duncan法进行多重比较(P<0.05),并对处理结果进行双因素(ANOVAs)分析。参照李京蓉等[18]的方法对4种植物采用隶属函数法进行综合评价。
2 结果与分析
2.1 光强对渗透调节物含量的影响
不同的光强对4种鸭跖草科植物的SS含量影响不同(图1)。CP的SS含量在L0~L3光强间无显著差异,而在L4光强下显著降低(P<0.05),TF的SS含量随光强的降低显著降低(P<0.05),与L0相比,在L4条件下降低了58.91%,而TV的SS含量在L0~L2光强间无显著差异,但均显著高于L3和L4条件。TZ的SS含量随光强的降低呈先升高后降低的趋势,在L3条件下上升到最高值,在L4条件下降到最低值,与最高值相比显著降低了22.06%(P<0.05);
在不同的光强下,4种植物的SP含量呈相同的变化趋势(图1),均随光强的降低而显著降低(P<0.05)。与L0条件相比,CP,TF,TZ和TV在L4条件的SP含量分别降低了48.68%,65.31%,70.28%,57.39%,其中TZ的降低幅度最大。

图1 不同光强对渗透调节物含量的影响
4种植物的Pro含量随光强的降低呈先降低后增加的趋势,且在L0和L4条件下的Pro含量均高于L1,L2和L3光强条件,但TF和TV在各处理间的差异均不显著(图1)。CP在L0条件下达到峰值,是最低值(L3条件下)的1.06倍,TZ在L4条件下达到峰值,是最低值(L1条件下)的1.11倍。
2.2 光强对MDA含量的影响
不同的光强下,4种植物的MDA含量的变化趋势相同,均随光强的降低而显著降低(P<0.05)(图2)。与L0相比,CP,TF,TZ和TV在L4条件的MDA含量分别降低了42.60%,36.87%,73.65%,43.44%,且TZ和TV在L0~L3条件下的MDA含量高于CP和TF,这表明强光条件导致4种植物产生了膜脂过氧化反应,且TZ和TV的膜系统损伤更大。
2.3 光强对H2O2和含量的影响

图2 不同光强对丙二醛含量的影响

图3 不同光强对H2O2和的含量影响

图4 不同光强对叶片H2O2(DAB染色检验)和(NBT染色检验)的影响
2.4 光强对抗氧化酶活性的影响
随光强的降低,CP和TV的SOD活性均呈“高-低-高”的变化趋势(图5),在L0条件下活性最高,且都在L3条件下降到最低值,与L0相比分别显著降低了13.86%,17.93%(P<0.05);TF在L0和L1条件下的SOD活性显著高于L2~L4条件(P<0.05),TZ在L0条件下的SOD活性显著高于L1~L4条件(P<0.05)。
4种植物的CAT活性在不同的光强下呈不同的变化趋势(图5)。CP的CAT活性在各处理间无显著差异,TF和TV的CAT活性随光强的降低呈“高-低-高”的变化趋势,其中,TF在L3条件下降到最低值,与L0相比,显著将低了60.09% (P<0.05),TV在L2条件下降到最低值,与L0相比,显著降低了35.49%;TZ的CAT活性与光强正相关,在L4条件下降到最低,与L0相比,显著降低了40.63%。
4种植物的POD活性随光强的降低显著降低(P<0.05)(图5),其中CP在L4条件下降到最低值,显著降低了50.29%(P<0.05);TF,TZ和TV均在L3条件下降到最低值,与L0相比分别显著降低了70.39%,56.80%,60.15%。
2.5 光强对叶肉细胞超微结构的影响
不同的光强对4种植物叶肉细胞的超微结构形态产生了不同程度的影响(图6、图7、图8)。CP在L0条件下,细胞结构产生了严重的损伤,细胞膜解体并与细胞壁分离;叶绿体发生严重的形变,基粒片层和基质片层蜷曲,排列松散且片层间隙大,细胞器无规律的散布在细胞中,淀粉粒体积大,几乎占据整个叶绿体。在L1~L4条件下,细胞质壁结构完好并结合紧密,叶绿体无溶解破碎现象,各片层结构排列紧密,呈纺锤形紧密贴合在细胞壁上,无堆叠挤压的情况出现,淀粉粒体积也逐渐变小。不同的光环境对CP的叶绿体、淀粉粒、嗜锇颗粒数量产生了显著的影响。叶绿体个数随着光强的减小而显著增加(P<0.05),与L0相比,在L4条件下增加了39.57%;淀粉粒个数在L0条件下达到峰值,显著高于其他光环境(P<0.05),是最低值L4条件的3.47倍;嗜锇颗粒数量随光强的降低呈先降低后增加的趋势(图8),在L3条件下数量最少;线粒体个数在不同的光环境下无显著差异。

图5 不同光强对抗氧化酶的影响
TF的细胞质壁结构在5种不同光强下结合紧密,界限清晰。在L0条件下,叶绿体部分解体,多个叠合分布在细胞壁边缘,淀粉粒体积大,占据整个叶绿体结构的3/4左右;随着光强的降低,淀粉粒体积逐渐变小,叶绿体也由宽圆形变为长纺锤形,单个独立分布在细胞壁边缘,但在L4条件下,部分叶绿体也发生了形变,虽然叶绿体在不同光环境下产生了不同的变化,但基质片层和基粒片层始终排列规则紧密。不同的光强对叶绿体、淀粉粒、线粒体和嗜锇颗粒数量产生了显著的影响(P<0.05)。L0条件下的叶绿体数量显著低于其他光环境(P<0.05),在L3条件下达到峰值,而L0条件下的淀粉粒个数又显著高于其他光照条件,其中淀粉粒个数在L3条件下降到最小值,嗜锇颗粒随光强的降低而显著降低(P<0.05),在L3条件下数量最少,仅为L0条件下的34.78%,L1条件下的线粒体数量显著高于其他光条件(P<0.05)。
TZ的细胞质壁结构在L0和L1条件下出现了严重的损坏,与细胞质界限不清,部分细胞膜解体,在L2,L3和L4条件下结构完好。L0和L1条件下的叶绿体结构破损,质膜溶解,部分基质片层和基粒片层结构出现解体现象,呈长圆形游离在细胞壁边缘,淀粉粒体积大且出现空泡现象,在L2~L4条件下,叶绿体结构完好,片层结构紧密,紧贴细胞壁分布,淀粉粒体积也逐渐变小,在L4条件下消失。不同的光强对TZ的叶绿体、淀粉粒和嗜锇颗粒数量产生了显著的影响(P<0.05),与L0相比,在L4条件下叶绿体数量增加了30.73%,淀粉粒数量降低了79.15%,嗜锇颗粒数量随光强的降低而显著降低,在L0条件下数量最多,在L2条件下最少,显著降低了58.58%,线粒体数量在各处理下无显著变化。
TV的细胞质壁结构在L0条件下发生了质壁分离的现象,但与细胞质界限清晰,在其他光环境下质壁结构完好。叶绿体在L0和L1条件下质膜解体,变为圆形,出现了堆叠现象,少量片层结构出现卷曲现象,但排列仍较为紧密,整个叶绿体被淀粉粒充斥。在L2~L4条件下,叶绿体结构完好,基质片层和基粒片层排列规则紧密,紧贴细胞壁分布,但仍有部分叶绿体发生形变现象,淀粉粒体积逐渐变小。不同的光强对TV的叶绿体、线粒体和嗜锇颗粒数量产生了显著的影响(P<0.05),叶绿体数量在L2条件下最多,在L4条件下最少。线粒体数量在L0条件下最少,显著低于L1和L2条件(P<0.05),而其他处理间无显著差异。嗜锇颗粒数量随光强的降低显著降低(P<0.05),在L3条件下最少,与L0相比,显著降低了48.42%。

图7 不同光强对叶绿体结构的影响(12000×)
2.6 物种和光强对不同指标的双因素方差分析
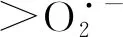

图8 不同光强对叶肉细胞指标的影响

表2 物种和光强对不同指标的双因素方差分析
2.7 4种植物的适光性综合评价
通过隶属函数法对4种鸭跖草科植物的适光能力进行了综合评价(表3),结果表明适光性强弱的的顺序为:吊竹梅>紫鸭跖草>‘花叶’水竹草>‘绿叶’水竹草。

表3 不同光强下4种植物生理指标的隶属函数值
3 讨论
渗透调节物质在植物抵抗非生物胁迫时发挥着举足轻重的作用。当产生的活性氧超过抗氧化物清除能力时,细胞质膜会发生严重的膜脂过氧化反应,此时植物细胞膜的通透性就会变大,导致细胞质外渗,造成植物生理代谢紊乱[19],此时植物就会通过调整SS,SP,Pro等渗透调节物质的含量来减轻逆境胁迫引起的细胞失水,以维持细胞渗透压平衡,从而降低逆境伤害的程度[20]。SS和SP既是重要的营养物质,又参与植物细胞的渗透调节过程[21],其中SS还可清除液泡和叶绿体中的ROS[22],可间接激活抗氧化酶系统。Pro被证明是一种有效的自由基清除剂,对稳定原生质胶体、细胞代谢过程、维持渗透调节平衡等方面起重要作用[23-24]。本研究发现‘花叶’水竹草和‘绿叶’水竹草的SS含量和SP含量均随光强的降低而显著降低,这表明这两种植物在强光胁迫下均能通过增加SS含量和SP含量,以清除液泡和叶绿体中的ROS,并通过碳代谢途径和氮代谢途径协同作用进行渗透调节,维持细胞膨压;而紫鸭跖草和吊竹梅的SS含量随着光强的降低呈先升高后降低的趋势,SP含量则随光强的降低而显著降低,这表明这2种植物在强光胁迫下氮代谢途径对渗透调节的贡献比碳代谢途径大,而在75%~25%自然光强中,碳代谢途径对渗透调节的贡献大于氮代谢途径。4种植物的SS含量和SP含量均在L4条件下降到最低,这可能是由于光强不足,叶片的光合作用急剧下降,SS和SP合成受阻,最终会导致植物的营养物质短缺,对生长造成不利影响。‘花叶’水竹草和‘绿叶’水竹草的Pro含量在各处理间无显著差异,这表明这2种植物的Pro含量稳定,始终保持相对较高的含量,清除植物细胞内的自由基,并抵消光强因素对细胞渗透调节的影响,而紫鸭跖草和吊竹梅的Pro含量随光强的降低呈先降后增的趋势,这表明Pro在极端弱光中仍能很好的发挥渗透调节的作用。

4 结论

猜你喜欢
农业技术与装备(2022年6期)2022-08-17
少儿科技(2019年9期)2019-09-10
债券(2019年1期)2019-09-05
西安工程大学学报(2016年6期)2017-01-15
中国骨科临床与基础研究杂志(2017年5期)2017-01-12
广西林业科学(2016年1期)2016-03-20
中国粮油学报(2016年1期)2016-02-06
食品工业科技(2014年23期)2014-03-11
中国烟草学报(2012年2期)2012-04-09
植物营养与肥料学报(2010年2期)2010-10-24
