
南欧蒜有性繁殖株系表型性状的比较分析
2021-03-29 05:43:46青李锡香宋江萍张晓辉武亚红都真真胡俊玲马龙传王海平
中国蔬菜 2021年3期
赵 青李锡香宋江萍张晓辉武亚红都真真胡俊玲马龙传王海平*
(1 中国农业科学院蔬菜花卉研究所,农业农村部园艺作物生物学与种质创制重点实验室,北京 100081;2 山东省济宁市金乡县大蒜工程技术研究中心,山东济宁 272200)
南欧蒜(Allium ampeloprasumL.)是一种大蒜近缘种资源(Tchórzewska et al.,2015),原产地在地中海地区,包括南欧、北非和西亚,后来引进到北美、南美、澳大利亚及其他地区(Aedo et al.,2013)。南欧蒜产量高(谷卫刚 等,2006),能量低,是膳食纤维和锌的良好来源,主要的脂肪酸是亚油酸,具有比常规蔬菜高比例的多不饱和脂肪酸(García-Herrera et al.,2014),其精油主要成分为甲基丙基二硫化物、樟脑、二甲基呋喃等,具有减少肠癌发生、降低血液胆固醇含量等重要的药用价值(Gantait et al.,2010;Monemi et al.,2014),受到了多个国家和地区特别是欧洲国家和地区的重视(Clercq et al.,1999;Monemi et al.,2014),在我国也逐渐推广种植,具有很好的产业前景。
前人对植物生长动态观察发现,植株在生长阶段的发育速度差异明显,反映出植株之间的生长优劣及营养分配对育性的影响。全杜娟等(2012)对角果藜的生长动态观察发现在株高增加迅速的阶段,果实积累加快。王裕欣等(2003)对豫芝11号芝麻生长动态观察发现,茎秆、叶面积系数在生长发育后期增长缓慢,蒴果质量积累加快,光合产物分配倾向于生殖生长。胡根海等(2002)对北疆春大豆的生长动态观察发现,株高与叶面积均在结荚盛期达到最高值,而鼓粒期至成熟期,营养物质向荚粒运送,籽粒充实度增加。一直以来,普通大蒜的繁殖方式以无性繁殖为主,因普通大蒜花发育后期受营养生长竞争影响,生殖生长受阻(Koul &Gohil,1970),导致不能结实,无法对其进行生长动态观测。南欧蒜作为有性繁殖的大蒜近缘植物,具有明显的生殖生长特性。对大蒜近缘植物南欧蒜生长动态进行观察,了解营养生长和生殖生长的内在联系,可为普通大蒜育性机理研究及南欧蒜的开发利用提供参考。
1 材料与方法
1.1 试验材料
试验材料为国家无性繁殖及多年生蔬菜种质资源圃保存的11 个南欧蒜株系,由播种南欧蒜8N540 的11 个实生种子,再由鳞茎分瓣方式繁殖而来,编号分别为8N540-1、8N540-2、8N540-3、8N540-4、8N540-5、8N540-7、8N540-8、8N540-9、8N540-10、8N540-11、8N540-12。
1.2 试验方法
试验材料于2017 年10 月中旬种植在中国农业科学院廊坊万庄国际高新技术产业园区,每个株系种植1 个小区,小区面积4.5 m2,以行距20 cm、株距15 cm 的密度播种。由于有性繁殖种子数量限制,未设重复。管理按常规进行。大蒜生长发育阶段分为萌芽期、幼苗期、花芽和鳞芽分化期、蒜薹伸长期、鳞茎膨大期、休眠期(张愚和毛啓明,1963),不同指标的调查方法依据《大蒜种质资源描述规范和数据标准》(李锡香和朱德蔚,2006)。2018 年3 月14 日至6 月13 日按不同发育时期调查后代株系的形态性状,如株高、开展度、叶长、叶宽、单株叶片数,每个株系调查10 株。在蒜薹伸长期调查地上假茎高、地上假茎粗2 个数量性状。在幼苗期和蒜薹伸长期测定植株生理性状,使用叶绿素测定仪选择最长叶测定叶片叶绿素的相对含量SPAD 值。在蒜薹伸长期调查后代株系的7 个质量性状,包括株型、叶片挺直度、叶横切面、叶色、叶面蜡粉、叶鞘色、假茎横切面,每个株系调查10 株。选择8N540-8 株系一半植株自交授粉,一半植株开放授粉,其他株系开放授粉,统计株系结实率。
1.3 数据处理与分析
利用Microsoft Excel 2016 软件对试验数据进行整理,对不同时期的株高、开展度、叶长、叶宽、单株叶片数进行变化规律分析,并根据叶面积模拟方程(谷卫刚,2003)y=2.004 7X1+9.122 1X2-52.933 4(X1:叶宽,X2:叶长)计算叶面积。利用SPSS 26.0 软件对数量性状进行方差分析、正态性检测及相关性分析,对于正态分布的数量性状,根据(X-1.281 8S)、(X-0.542 6S)、(X+0.542 6S)和(X+1.281 8S)(S是标准差)4 分点原则并依据孙升(1999)的分级方法将数量性状分为1、3、5、7、9 等5 个等级,对各性状进行数量分级并整理,同时将质量性状数量化,通过公式:H′=-∑PilnPi(其中i为某一性状的分级,Pi为第i等级样品的频率,即在群体中所占的频率)计算各性状的Shannon-Weiner 多样性指数(H′)。
2 结果与分析
2.1 南欧蒜不同株系株高、开展度的动态变化分析
株高、开展度是影响大蒜株型的重要农艺性状。结果表明,南欧蒜所有株系在整体上呈现相似的株高变化规律(图1-A)。3 月14 日至4 月20 日,11 个株系的株高不断增长,其中4 月16 日至4 月20 日,植株生长速度最快,处于旺盛生长期,植株以营养生长为主;4 月20 日至5 月17 日,株高的增长放缓,植株处于营养生长和生殖生长并重的时期;5 月17 日至6 月5 日,大部分植株株高的增长更为缓慢,此期植株已经抽薹,处于花茎伸长期,植株以生殖生长为主;6 月5 日至6 月10 日,部分植株株高降低,主要是由于这些植株叶片开始下垂。在11 个株系中,8N540-11 在绝大部分生育期的株高均超过其他株系,且其在3 月14 日至4月2 日、6 月5 日至6 月10 日的生长速度均超过其他株系(图1-A)。
开展度与株高有相似的变化规律,但是并不完全相同,3 月14 日至4 月20 日,植株开展度不断增长,其中4 月16 日至4 月20 日之间增长速度最快,处于旺盛生长期;4 月20 日至5 月17 日,植株开展度变化不大;5 月17 日至6 月5 日,部分株系的开展度较株高更早出现下降趋势,说明叶片下垂对开展度影响更大;6 月5 日至6 月10 日,部分株系植株的开展度增长。在整个植株发育过程中,8N540-11 株系的开展度远远超过其他株系,且其在4 月20 日至5 月17 日的生长速度超过其他株系,在5 月17 日至6 月5 日开展度的下降速度快(图1-B)。
从整体上来看,11 个不同株系的株高与开展度在营养生长为主的阶段均表现出缓慢生长、快速生长、缓慢生长的发育规律,其中8N540-11 株系的株高、开展度变化速度均大于其他株系。
2.2 南欧蒜不同株系叶片的动态变化分析
在南欧蒜生长发育过程中,所有株系均呈现类似的叶长变化规律(图2-A)。从3 月14 日至4 月16 日,植株叶长不断增长;4 月16 日至4 月20 日,植株叶长的增长速度最快,达到旺盛生长期;4 月20 日至5 月17 日,植株叶长的增长速度减缓;5月17 日至6 月10 日,植株叶长的增长速度更为缓慢。在整个生长发育过程中,8N540-11 株系的叶长均超过其他株系,且其在4 月9 日至4 月16 日的叶长增长速度达到最大值(图2-A)。所有株系的叶宽都有类似的变化规律,3 月14 日至5 月17日,植株叶宽不断增长;5 月17 日后,植株叶宽不断下降,此时植株已经抽薹,处于生殖生长期,叶片聚拢。在整个生长发育过程中,8N540-11 株系的叶宽均超过其他株系(图2-B)。大部分株系的单株叶片数在4 月20 日前不断增加,此后不断减少,表明植株生殖生长吸收了部分营养物质,部分叶片开始脱落。在整个生育期,8N540-11 株系的单株叶片数均超过其他株系(图2-C)。依照谷卫刚(2003)的叶面积模拟方程计算获得的叶面积与叶长呈现相同的变化规律,表明叶面积主要与叶长有关(图2-D)。
从整体上看,11 个不同株系的叶长、叶面积在营养生长为主的阶段均表现缓慢生长、快速生长、缓慢生长的发育规律。在整个生育期内,8N540-11 株系的叶长、叶宽、单株叶片数、叶面积均超过其他株系。
2.3 南欧蒜不同株系叶绿素相对含量的差异
通过叶绿素测定仪测定幼苗期与蒜薹伸长期不同株系叶片的叶绿素相对含量。结果表明(图3),幼苗期叶绿素相对含量最高的是8N540-2,最低的是8N540-10。蒜薹伸长期叶绿素相对含量最高的是8N540-11,最低的是8N540-3。在这2 个时期8N540-3、8N540-10、8N540-11 的叶绿素相对含量差异显著。
2.4 南欧蒜不同株系数量性状的变异
对南欧蒜8 个数量性状的变异情况进行分析,结果表明(表1),南欧蒜后代株系的形态性状,包括株高、开展度、叶长、叶宽、单株叶片数、地上假茎高、地上假茎粗差异明显,变异系数分布范围在19.56%~27.35%,平均为23.55%,其中开展度的变异系数最大,叶长的变异系数最小。生理性状叶绿素相对含量的变异系数为24.87%,表明后代株系的叶绿素相对含量差异明显。各形态性状的多样性指数分布在0.60~1.04 之间,地上假茎粗、叶长的多样性指数较大,单株叶片数的多样性指数较小,而且变异系数大的性状,其多样性指数不一定大。生理性状叶绿素相对含量的多样性指数达1.21,高于所有形态性状。
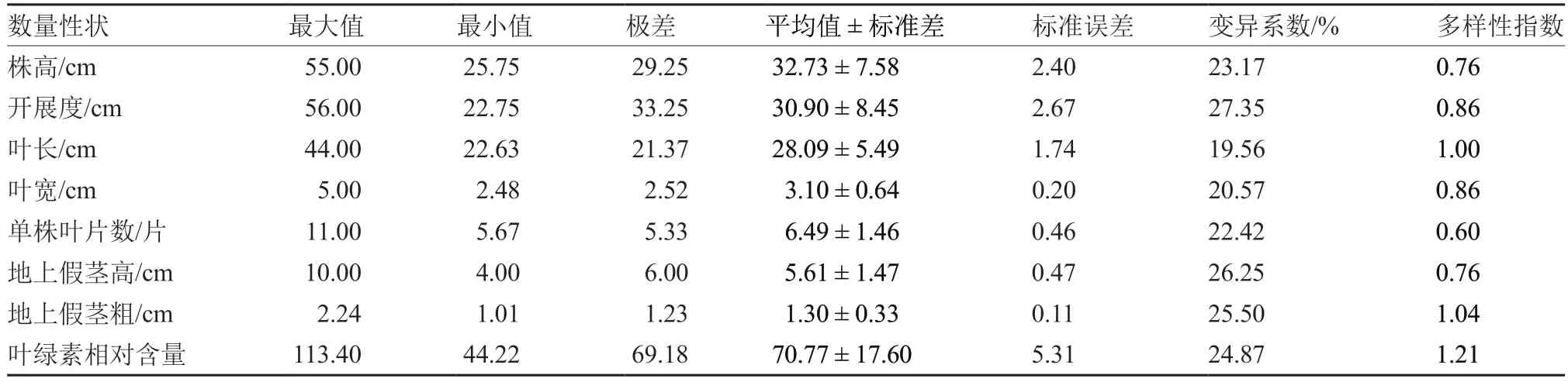
表1 南欧蒜不同株系8 个数量性状的变异情况
2.5 南欧蒜不同株系质量性状的表现
对南欧蒜不同株系质量性状进行分析,株型表现直立与半直立2 种类型,叶面蜡粉表现中、多2种类型,假茎横切面表现圆形与椭圆形2 种类型,叶片挺直度、叶横切面、叶色以及叶鞘色表现一致,分别是叶片挺直、叶横切面为“V”型、叶色绿、叶鞘绿白色(表2)。质量性状的多样性指数反映了性状在不同级别上的分布情况,株型、叶面蜡粉、假茎横切面的多样性指数在0~0.36 之间,表明后代株系的株型、叶面蜡粉、假茎横切面具有丰富的遗传多样性。
2.6 南欧蒜不同株系的结实率
选择8N540-8 株系一半植株自交授粉,一半植株开放授粉,其余株系开放授粉,观察其结实情况(表3)。8N540-8 株系采用自交授粉方式的结实率高于开放授粉,说明人工辅助授粉能提高南欧蒜结实率。
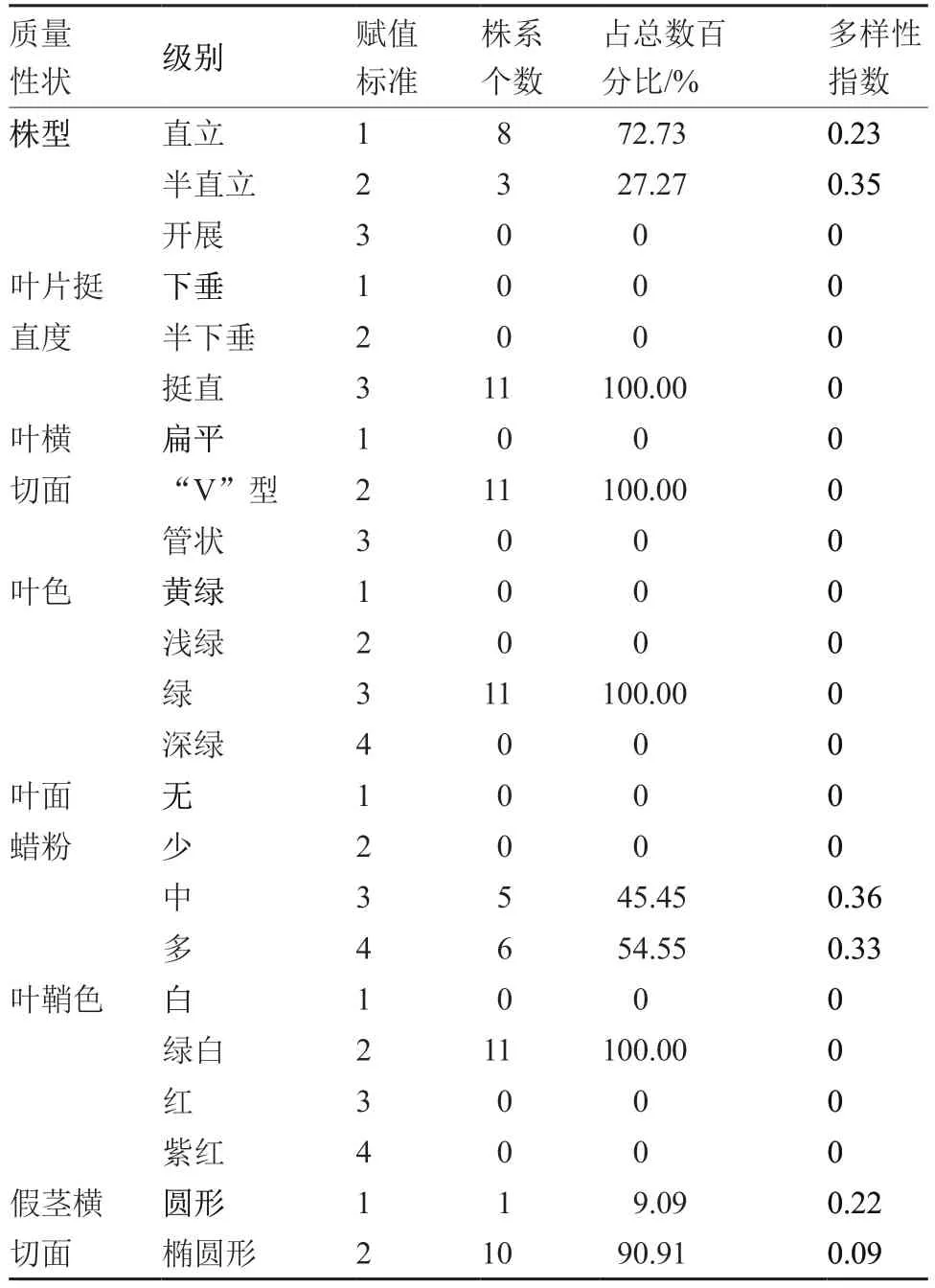
表2 南欧蒜不同株系7 个质量性状的多样性
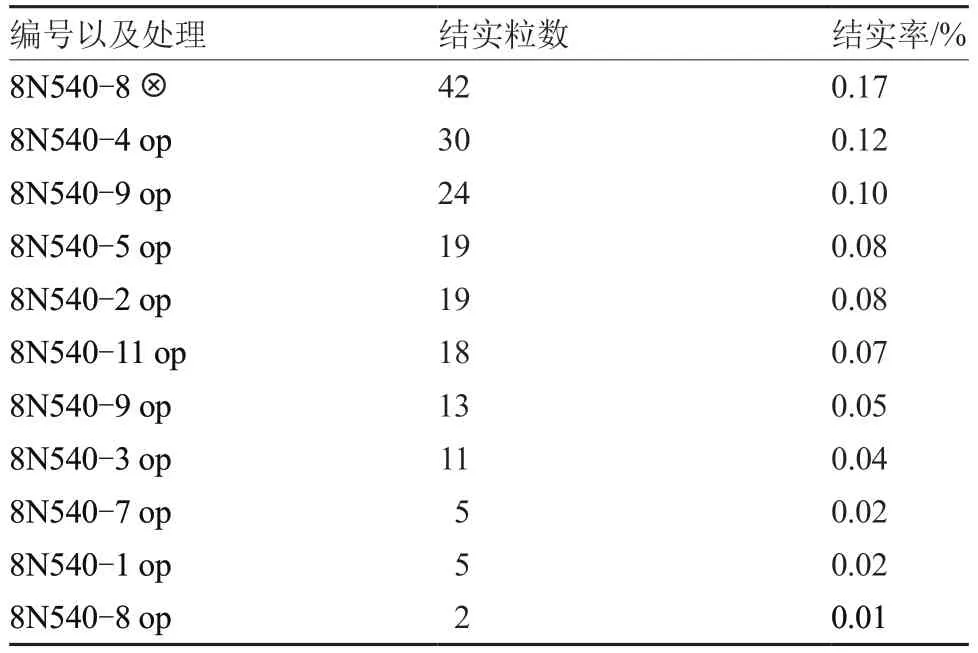
表3 南欧蒜不同株系的结实率
3 结论与讨论
本试验发现南欧蒜不同株系的株高、开展度、叶长等形态性状均大体呈现出缓慢生长、快速生长、缓慢生长的生长规律,在生长末期还会出现株高、开展度下降。这与刘秀慧(2017)对5 个栽培大蒜品种生长动态观测的结果基本一致。在4 月16 日至4 月20 日之间,11 个南欧蒜株系处于旺盛生长期,此期是南欧蒜生长管理的关键时期,这比刘秀慧(2017)发现的5 个栽培大蒜品种的旺盛生长期早7 d 左右,南欧蒜的生长发育速度可能较普通大蒜快,为其生殖生长奠定了足够的能量储备。本试验中南欧蒜的生长周期约290 d,比普通大蒜长15 d 左右,这与谷卫刚(2003)对南欧蒜的生长发育时期研究结论相一致。本试验还发现部分南欧蒜株系的开展度下降之后还会出现上升的状况,可能是因为鳞茎膨大完成后,叶片尚未失绿,营养物质被运往蒜薹,促进了开展度的增长。
叶绿素含量是影响大蒜光合效率的重要因素,其含量高低直接影响大蒜光合能力的强弱(张亚娟等,2016),本试验对南欧蒜叶绿素相对含量进行测定后发现,与其他株系相比,8N540-11 株系在蒜薹伸长期的叶绿素相对含量最高,表明8N540-11 株系在蒜薹伸长期的生长发育状况可能优于其他株系。
本试验通过对数量性状和质量性状变化研究发现南欧蒜后代株系数量性状变异较大,这为挑选优良的株系类型奠定了基础。在南欧蒜后代株系中,8N540-11 株系的株高、开展度、叶长、叶宽、单株叶片数、叶面积均超过其他株系,且其结实率中等,综合性状表现较好,可见种子繁殖南欧蒜可以创造生长势强的株系。
总之,本试验发现南欧蒜有性繁殖后代的生长发育及性状表现有很大差异,为南欧蒜种质创新提供了材料基础,特别是8N540-11 株系表现出明显的优势,生长速度快,具有育种的潜在价值。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17 12:39:32
云南师范大学学报(自然科学版)(2022年2期)2022-03-29 02:58:16
新疆农业科学(2020年1期)2020-02-14 03:22:46
亚热带植物科学(2019年2期)2019-07-22 13:16:12
四川农业科技(2019年5期)2019-07-01 09:46:46
湘潮(上半月)(2019年10期)2019-05-22 06:08:50
江苏农业科学(2016年10期)2017-02-05 14:45:09
浙江柑橘(2016年1期)2016-03-11 20:12:31
水电站机电技术(2016年1期)2016-02-28 14:21:45
戏剧艺术(上海戏剧学院学报)(2016年5期)2016-01-23 02:36:41
