
菇木林目标树择伐林窗的形成对土壤养分含量的影响
2021-03-17 06:43:32
中南林业科技大学学报 2021年2期
(中国林业科学研究院 亚热带林业研究所,浙江 杭州 311400)
森林生态系统易受自然或人为因素干扰而形成林窗[1-2],林窗的尺度往往较小,是森林生态系统中普遍存在的现象。林窗也是森林群落演替的重要驱动要素,对森林生态系统的结构和功能有着显著的影响[3-4]。林窗的存在使得林窗内微环境发生改变[5],非生物环境如林窗内太阳辐射和光照强度[6]、空气温度和湿度、土壤温度和湿度[7]、土壤pH 值[8]以及生物环境如物种更新[9-11]、植物多样性[12-13]、土壤微生物和土壤动物等[14-16],林窗内生物环境、非生物环境都与非林窗区域有着明显的不同,从而影响林窗内凋落物分解和养分释放[17]及森林生态系统养分循环和能量流动[18],林窗的形成有利于森林群落演替和生态系统发展[19-20]。
菇木林是指以生产剁花法香菇为目的的林分类型(不同于一般意义上的用于生产香菇培养料的林分)[21],剁花法香菇栽培是一种传统的香菇生产技术,有着800 多年的历史,通过在林地原地砍倒适合做香菇的阔叶树木(即菇木林里面的林木),并在倒木上剁出疤痕,利用自然孢子传播接种栽培香菇。菇木林采伐通常采用“伐大留小”的异龄林择伐技术,主要是将胸径20 cm 以上的菇木按照一定的技术要求砍倒,砍伐后常常会在林内留下30~100 m2不等的人工林窗,而且伐倒的菇木(包括枝条和叶子)全部生物量留在原位腐烂分解,以保证林木养分的自我归还,是一种典型的人与自然和谐发展的森林经营技术,我国南方地区,特别是浙江、江西、福建、广东等省都有曾用作菇木林的常绿阔叶林[21]。目前关于林窗的研究局限于天然形成的林窗,而对于天然林人工择伐后形成的林窗则研究较少[2,4,13,22],特别是关于我国亚热带东南沿海地区天然林择伐后所有生物量留在原位地表形成的人工林窗的研究更为少见。菇木林人工形成的林窗土壤养分元素和土壤质量对菇木择伐响应如何,对于传统菇木林的科学经营有何指导意义等问题急需回答。本研究以菇木林人工择伐后,所有采伐物全部留在原位而形成的人工林窗为研究对象,研究菇木林目标树种择伐人工林窗对土壤养分元素含量的影响,揭示菇木择伐形成的林窗效应,为中国重要农业文化遗产——浙江省庆元香菇文化系统的培育、推广和优化管理提供科学依据。
1 研究地概况与研究方法
1.1 研究地概况
研究地设在中国重要农业文化遗产——浙江省庆元香菇文化系统内(地理位置为118°50′~119°30′E,27°25′~27°51′N),气候属于亚热带季风气候区,温暖湿润,冬暖夏凉,年均气温17.4℃,年均降水量1 760 mm,无霜期245 d。研究地地带性植被为亚热带常绿阔叶林,乔木植物以红栲Castanopsis hystrix、木荷Schima superba、乌槠Castanopsi sclerophylla、甜槠Castanopsi eyrei、米槠Castanopsi carlesii为主,林下灌木植物以格药柃Eurya muricata、朱砂根Ardisia crenanta、四川山矾Symplocos setchuensis为主,草本植物有蕨Pteridium aquilinum、芒萁Dicranopteris pedata等。研究样地土壤为红黄壤,土层深厚,有机质含量高,土体疏松,腐殖质层厚。
1.2 研究方法
1.2.1 试验设计和测定方法
试验地林木平均胸径为15.2 cm,平均树高为12.6 m,密度为1 600 株·hm-2,林龄为成熟林,封山育林60 a 以上,总面积125 hm2。2012年12月,选择适宜培育剁花香菇的间隔一定距离的红栲树5株(菇木平均胸径23.2 cm,相当于林窗单位面积形成的树干分解物为16.14 t·hm-2)[23],利用地形,从下而上,定向将菇木砍倒,各形成5 个人工林窗;树梢向下坡,树基朝上坡放置树干,去其大部分枝杈,留下树干及少量枝杈;接着根据菇木的情况用斧头剁出不同深度和斜度的疤痕,同时用砍下的树枝和树叶(相当于林窗单位面积内形成的枝条和叶子分解物的数量为14.07 t·hm-2,因此留在林窗内腐烂的树干、枝条和叶子合计为30.11 t·hm-2)等覆盖菇木,防止烈日暴晒,形成的林窗面积为80 m2左右。剁花5 a 后菇木基本不再生产香菇,树干也接近腐烂。2018年8月,于砍伐后形成的每一个林窗内的南北轴和东西轴上,各间隔2~3 m设立5 个土壤采集样点,去除地表枯落物后,用内径为5 cm 的土钻,采集0~20、20~40 和40~60 cm 土层的土壤样品,每一个林窗同一土层的土壤样品均匀混合。同时在每一个林窗外10 m范围内设立对照林地各1 个,样地大小为20 m×20 m,每一个样地按照“S”型取样法,采用内径为5 cm 的土钻采集0~20、20~40、40~60 cm土层土壤,同一土层的土壤样品充分混合。土壤样品捡出植物根系、残体和碎石,自然风干后过2 mm 筛,干燥保存,用于测定土壤有机碳和土壤养分元素含量。选择对照林地时,尽量保持其与菇木林地的植被、坡度、坡向、海拔等自然地理条件基本一致,以保证试验的可比性。
土壤有机碳(TOC)的测定采用重铬酸钾外加热法[24]。土壤养分的测定采用常规方法[25]:全氮(TN)采用凯氏定氮法测定;水解氮(AN)采用碱解扩散法测定;速效钾(AK)采用乙酸浸提法测定;有效磷(AP)采用碳酸氢钠法测定;土壤交换性钙(Ca)和交换性镁(Mg)的测定采用原子吸收分光光度法;土壤pH 值采用电位法测定。
1.2.2 统计分析
采用SPSS16.0 统计软件包中的t 检验分析法(Independent-samples T Test)比较养分元素的变化情况,采用SPSS16.0 统计软件包中的主成分分析方法(Principal components analysis)对土壤养分元素进行主成分分析,并对综合评价值F进行排序(F=∑yi得分×yi贡献率,公式中yi分别代表第i个主成分的得分和贡献率,i=1,2,3,…),根据F值的大小对菇木林窗和对照林分进行土壤肥力评价[26])。
2 结果与分析
2.1 菇木林窗对土壤pH 值及C、N 含量和比值的影响
菇木林窗和对照林土壤pH 值及C、N 含量和比值见表1。由表1可知,与对照林地相比,林窗0~20、40~60 cm 土层土壤pH 值分别下降了2.79%和0.56%,20~40 cm 土层土壤pH 值则增加了0.94%,但差异均不显著。与对照林地相比,0~20 cm 土层土壤有机碳含量增加了25.55%,但差异不显著;而20~40、40~60 cm 土层林窗土壤有机碳含量分别下降了43.5%、54.68%,且差异达到了显著水平。与对照相比,林窗0~20 cm土层全氮含量增加了13.68%,差异不显著;而20~40、40~60 cm 土层的全氮含量分别下降了37.31%和44.43%,且差异达到显著水平。与对照相比,林窗0~20 cm 土层土壤C∶N 提高了10.04%,但差异不显著;20~40、40~60 cm 土层的C∶N 分别下降了9.05%、19.65%,且差异都达到了显著水平。
2.2 菇木林林窗对土壤速效养分含量的影响
菇木林窗和对照林土壤水解性氮、有效磷和速效钾含量见表2。由表2可知,与对照相比,择伐后留在林窗的菇木及其枝叶的分解有利于0~20 cm土层水解性氮含量的增加,增加了42.34%,但差异不显著;20~40、40~60 cm 土层,水解性氮含量下降了23.49%和10.28%,且20~40 cm 土层差异显著。与对照相比,菇木林窗有效磷的含量同水解氮的变化趋势一致,0~20 cm 土层增加了15.32%,但差异不显著;20~40、40~60 cm 土层分别下降了11.06%和34.47%,且40~60 cm土层差异显著。与对照相比,菇木林窗0~20、20~40、40~60 cm 土层速效钾含量分别下降了28.02%、35.08%和0.25%,其中20~40 cm 土层差异达到显著水平。
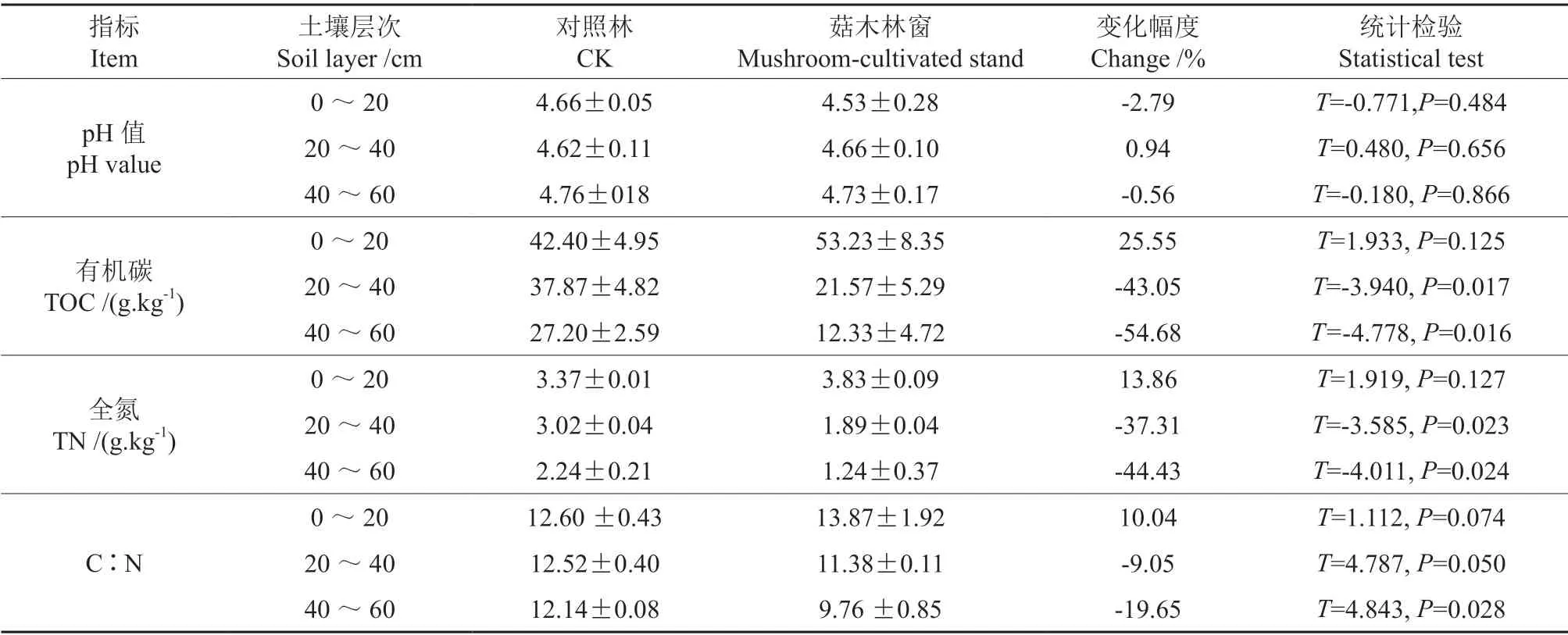
表1 菇木林窗和对照林土壤pH 值及C、N 含量和比值的比较†Table 1 Soil pH,TOC,TN and C/ N ratio of soil in mushroom-cultivated stand gap and CK plots
2.3 菇木林林窗对土壤交换性钙、交换性镁含量的影响
与对照相比,林窗0~20、20~40、40~60 cm 土层交换性钙含量分别增加了83.16%、35.78% 和42.00%,且在0~20 cm 土层差异达到显著水平;与对照相比,林窗0~20、20~40、40~60 cm 土层交换性镁含量分别下降了19.26%、20.71%和7.18%,但是差异均不显著(表3)。
2.4 菇木林窗和对照林土壤肥力的主成分分析
对表征土壤肥力的因子(0~20、20~40、40~60 cm 土壤pH 值、有机碳、全氮、速效氮、有效磷和速效钾、交换性钙和交换性镁含量均值建立数据矩阵)进行主成分分析,结果(表4)表明,菇木林林窗两个主成分(y1,y2)的方差贡献率分别为83.02%、10.70%,累计贡献率为93.72%;对照林两个主成分(y1,y2)的方差贡献率分别为75.39%、18.38%,累计贡献率为93.77%。因此,两个主成分足以代替原始因子所代表的主要信息。由综合评价值F=∑yi得分×yi贡献率(i=1,2),求得菇木林林窗和对照林的F得分值,分别为0.823 5、0.764 1,前者大于后者,菇木林林窗有利于土壤质量的提升。

表2 菇木林窗和对照林土壤水解性氮、有效磷和速效钾含量的比较Table 2 Comparison of soil AN,AP and AK between mushroom-cultivated stand gap and CK plots

表3 菇木林窗和对照林土壤交换性钙和交换性镁含量的比较Table 3 Comparison of soil exchangeable Ca and exchangeable Mg between mushroom-cultivated stand gap and CK plots
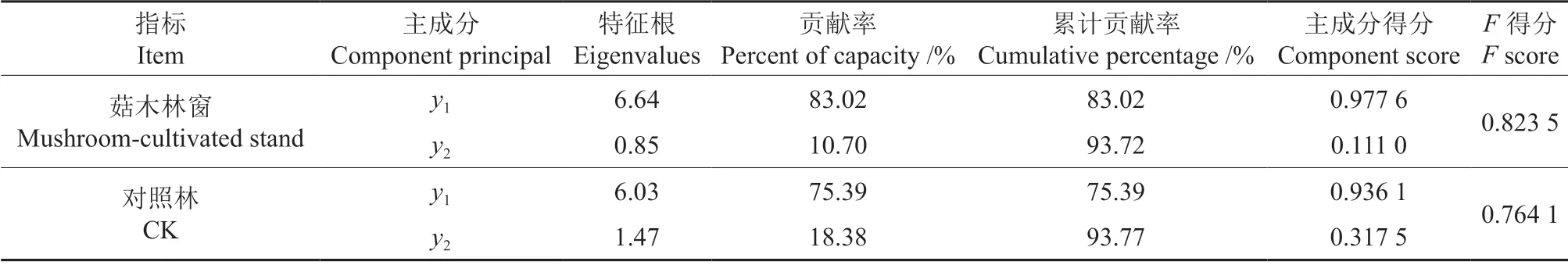
表4 菇木林窗和对照林土壤肥力比较Table 4 Principal component analysis of soil fertility in mushroom-cultivated stand gap and CK plots
3 结论与讨论
菇木形成的林窗改变了林内的光照条件和降雨条件,林窗内的温度和湿度发生相应的变化[27-28],影响着土壤微生物的活性[29],从而引起土壤养分元素含量的变化[30]。
菇木伐倒后,所有生物量原位返还到地表,林窗内地表枯落物数量(叶子和枝条)要较对照林分增加14.07 t·hm-2,加上菇木树干本身的腐烂分解,以及由于林窗内温度和湿度的改变导致微生物分解活性增强,导致林窗内0~20 cm 土层土壤有机碳含量增加[15];而林窗内20~40、40~60 cm 土层,由于有机质来源减少(林窗内植被减少,根系凋落物和分泌物减少,导致深层土壤中的有机质补充减少),且土壤矿物质化作用的增强[31],所以有机碳含量下降。周义贵等[32]研究了四川米亚罗地区云杉Picea asperata林林窗效应,结果也表明林窗的形成促使地表层土壤有机质含量的增加;张春雨等[33]研究认为,长白山阔叶红松Pinus koraiensis林林隙内土壤有机碳含量增加。而王清奎等[31]研究认为,林窗形成后,林窗内的环境条件发生显著的变化,土壤有机碳的矿化作用随之增强[34],土壤有机碳含量下降。
土壤中的全氮来源于土壤有机质,与土壤有机质密切相关,本研究土壤全氮的变化规律与土壤有机碳含量的变化规律相一致,林窗地表增加的凋落物的分解有利于0~20 cm 土层全氮和水解性氮含量的增加[35]。耿玉清等[36]和刘聪等[37]的研究结果也表明,林窗能提高A 层土壤的全氮和水解氮的含量;张春雨等[33]的研究也表明,林隙土壤全N 含量比林内提高了7.67%。但是,对于林窗内20~40、40~60 cm 土层而言,由于土壤矿化速率的增加,加上植物对养分元素吸收量的减少,林窗内土壤养分有效性可能会增大[8];林冠层降雨截留量的减少,淋溶作用增加导致养分流失也会增强[30],且由于植物根系分泌物的减少,会导致养分元素含量下降。菇木林窗改变了林内的光照强度,林窗内温度和湿度随之发生变化[7],酶的活性增强,促使地表凋落物的分解,释放大量酸性物质[8],所以降低了0~20 cm和40~60 cm 土壤pH 值,20~40 cm 土层土壤pH 数值则稍有增加。周义贵等[32]和张春雨等[33]的研究也表明,林窗促使土壤pH 值下降,但差异不显著。
耿玉清等[36]和刘聪等[37]的研究表明,林窗的形成可提高A 层土壤有效磷的量。但是对于深层土壤而言,由于土壤矿化速率增加,根系分泌物的减少和植物对养分元素吸收量的减少,增加了林窗内土壤养分有效性,因此,淋溶导致土壤有效磷、速效钾养分流失作用也倾向于增加[29],所以,本研究速效磷的含量在20~40、40~60 cm土层,速效钾在0~20、20~40、40~60 cm土层都呈现下降的趋势,张春雨等[33]也得出相似的研究结果。
一方面,由于菇木林窗凋落物分解释放养分和土壤矿物质化作用的增加[34,38],导致林窗土壤交换性钙和镁含量增加。另外一方面,由于林窗的形成,调节了降雨的分配比例,增加了林内穿透雨降雨量,导致林窗内土壤交换性钙、镁元素的流失。由于菇木林植物体内杆、枝、叶钙元素含量是菇木林植物体内杆、枝、叶镁元素含量的14.2 倍,林窗内枯落物分解后释放的钙元素量不仅仅弥补了钙的流失,而且利于土壤交换性钙的积累,因此导致林窗内土壤交换性钙、镁元素变化趋势不一致,所以导致本研究中林窗土壤的交换性钙元素含量高于对照,而交换性镁元素含量则相反。
采用主成分分析,对表征土壤肥力的8 个因子(土壤pH 值、有机碳、全氮、速效氮、有效磷、速效钾、交换性钙和交换性镁含量)进行了分析,综合评价表明,菇木林窗形成后有利于土壤肥力的提升,研究结果为菇木林经营提供了一定理论依据。
影响土壤质量的土壤属性很多,土壤的理化性质、生物属性都可能是最终的评价指标,本研究仅仅局限于从土壤养分和土壤pH 值等方面考虑菇木林窗形成后土壤质量的变化,且菇木林窗形成的年龄只有6 a,研究期限较短。今后继续以所设立的固定样地为研究平台,持续开展长期定位监测和研究,并且从生物因子(林下植被、土壤生物)和非生物因子(环境因子)等方面进行综合评价,为菇木林择伐和林窗管理提供更加科学的依据。
猜你喜欢
西南林业大学学报(2022年5期)2022-10-17 09:56:38
农业机械学报(2020年1期)2020-03-13 00:27:38
西部林业科学(2018年4期)2018-02-14 22:43:50
现代园艺(2017年21期)2018-01-03 06:41:42
绿色科技(2016年16期)2016-10-11 06:53:51
浙江农业学报(2016年7期)2016-06-15 20:30:08
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:16
现代农业(2016年4期)2016-02-28 18:42:07
环境科技(2015年2期)2015-11-08 12:11:30
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
