
不同木霉菌对寒地盐碱土壤玉米杂交种光合特性及活性氧代谢的影响
2021-03-11 07:56付健王玉凤张翼飞杨丽娟杨克军
黑龙江八一农垦大学学报 2021年1期
付健,王玉凤,张翼飞,杨丽娟,杨克军
(黑龙江八一农垦大学农学院/黑龙江省教育厅寒地作物种质改良与栽培重点实验室,大庆 163319)
玉米是我国重要的粮食作物之一,约占我国粮食总产量的40%,在保障国家粮食安全战略中具有重要地位,且玉米在我国未来粮食增产进程中将占主导地位。黑龙江春玉米区是我国玉米的主要产区,也是我国重要的商品粮基地,因此保障黑龙江地区春玉米产量的稳定对国家农业经济发展至关重要,同时黑龙江也是苏打盐碱地集中分布区之一,盐碱地面积2.882×106hm2,严重制约该地区粮食产量。
作物的光合作用在生长发育过程中具有非常重要的作用,作物通过光合作用可以产生大量的有机物,其中大约90%的有机物最终转化为作物籽粒产量。已有相关研究表明,作物的光合作用对盐碱胁迫的响应非常敏感,盐碱胁迫下,作物的光合速率显著下降,对作物产生明显的光合抑制作用,最终会导致作物产量严重降低,甚至导致作物死亡[1]。其中盐碱胁迫下,作物光合作用中光系统II(PSII)对胁迫表现最为敏感,主要由于盐碱胁迫导致水势下降从而引起渗透胁迫,最终导致光合电子传递链中过度还原,进而产生光氧化损伤[2]。在遭遇盐碱胁迫时,作物体内会受到非常大的影响,导致体内聚集大量的活性氧(ROS),而大量的活性氧聚集会引起作物体内细胞膜质过氧化,同时引发作物体内抗氧化酶发生改变来应对ROS 损伤[3],随着盐碱胁迫导致的过氧化反应逐渐增强,作物体内的抗氧化酶和非酶防御系统会产生明显的响应机制,从而促使作物能够有效的抵御盐碱胁迫导致的氧化损伤[4-6]。盐碱胁迫下,玉米植株会产生过量的ROS,从而改变了玉米植物体内活性氧自由基清除酶活性,导致细胞膜质过氧化,最终导致植株体内丙二醛(MDA)含量大量积累[7]。作物在生长过程中会受到很多环境胁迫的损害,因此,在长期应对由于环境胁迫导致的ROS 积累造成的损伤时,作物体内形成了比较完整的应对胁迫损害的防御机制,这套防御机制主要起到协调作物体内过量ROS 的清除作用,它对维持作物体内的氧化还原平衡具有非常重要的作用,而作物在利用防御机制低于损伤的同时,作物体内自身也会产生一定量的渗透调节物质,使作物体内细胞的渗透压维持正常水平,使细胞能够正常进行吸水生长,从而保证作物在环境胁迫下能够健康生长[8]。
木霉菌(Trichoderma)是一类重要的生防因子和促进植物生长菌。目前,木霉菌在全世界农业生产中被广泛的应用,且表现出良好的抗逆和促进生长的效果。相关研究结果发现,木霉菌能够有效的在植物根部进行定殖生长,并通过自身的生防机制对植物进行生长调节,从而提高植物应对非生物胁迫时的耐受能力,木霉菌对缓解植物应对盐碱、干旱等环境胁迫方面发挥了重要作用[9-10]。木霉菌对植物的促生长作用包括提高种子萌发、提高叶片和根系生物量、增强对养分吸收等[11-13]。前人在棉花上施用木霉菌后发现,木霉菌对棉花叶片的叶绿素含量具有非常明显的促进作用,从而促进了棉花叶片的光合作用[14],同时使植物的干物质积累显著提高[15-16],这说明光合作用对植物的物质积累和生长发育具有重要作用。研究发现,木霉菌处理下,植株体内的抗氧化防御系统相关参数均在木霉菌促进下显著提高,缓解了ROS 损伤,使MDA 积累量显著减少,使植株应对环境胁迫的能力显著提高[17]。有研究也表明,植物在施用木霉菌后,其体内的钙、钾等离子浓度显著增加,而钠离子浓度则显著减少,从而提高了植物应对盐碱胁迫的抗逆性[18]。木霉菌还通过参与一些光合作用相关的基因表达,来提高植株应对环境胁迫的能力,从而使植株光合能力增强[19]。目前,关于木霉菌与植物抗盐碱性的研究多数针对于蔬菜及林木类,在粮食作物上应用较少,且都是人工模拟的盐碱环境,但是寒地天然盐碱土的情况比较复杂,人工无法模拟,因此有关寒地天然盐碱土胁迫条件下,不同种属木霉菌对不同基因型玉米幼苗光合作用特性及活性氧代谢的影响尚未见报道。
研究分析盐碱土壤胁迫下,施用哈茨木霉(Trichoderma harzianum)和长枝木霉(Trichoderma longibrachiatum)对提高玉米耐盐碱性的作用效果,主要通过研究盐碱土壤胁迫下施用不同木霉菌对玉米幼苗的光合荧光特性、光合色素及活性氧代谢的影响,以阐明在逆境胁迫下不同种属木霉菌增强玉米光合作用的机理和调节植物盐碱应激反应有关的渗透压保护机制、ROS 的清除的改善,为木霉菌在黑龙江寒地盐碱土壤中的应用和推广提供科学依据。
1 材料与方法
1.1 试验材料与设计
试验于黑龙江八一农垦大学农学院省级重点实验室进行。供试品种为试验筛选的玉米杂交种:盐碱敏感型品种真金8 号(ZJ8)和耐盐碱品种江育417(JY417),人工进行种子一致性挑选,然后对挑选好的种子进行消毒处理,并放置于恒温培养箱中25 ℃黑暗催芽,在催芽结束后,选取长势一致的种子种植于宽和高分别为15 cm 和10 cm 的盆中,盆栽所用盐碱土取自黑龙江省大庆市,盐碱土壤pH 为9.30,盆栽土壤在室内阴凉干燥通风处自然阴干,过2 mm筛网,每盆放土700 g。供试木霉菌分别为哈茨木霉(Trichoderma harzianum) 和长枝木霉(Trichoderma longibrachiatum),以分别施用浓度为 1×109spores·L-1哈茨木霉(TH)、长枝木霉(TL)为试验处理,以不施用木霉菌为对照处理(Con),完全随机设计,5 次重复。其中不同木霉菌液处理按照每升土施用200 mL 木霉菌孢子悬浮液在栽苗后立即浇入,对照处理浇入等量无菌液水,玉米在植物生长室内生长到三叶一心时期进行样品采集。植物生长室培养条件为:昼夜温度为(25±2)℃(/20±2)℃,每天光照12 h,光强为1 000 μmol·m-·2s-1,湿度为 60%~80%[20]。盆栽土壤基础理化性质见表1。

表1 盆栽土壤理化性质Table 1 Physicochemical properties of the experimental soil
1.2 玉米幼苗样品采集及预处理
施用木霉菌后27 d,在不同处理下选取3 株玉米幼苗叶片进行光合作用参数和叶绿素荧光参数的测定,测定完成后将各处理的3 株玉米幼苗第二片展开叶进行采集用于测定叶片叶绿素含量,最后采集用于测定酶活性指标的叶片,用锡纸包好之后迅速放入液氮中冷冻,并保存于-80 ℃冰箱中。
1.3 指标测定
1.3.1 光合特性测定
采用 Li-6400 光合仪(Li-COR Inc,USA)对玉米幼苗的第二片展开叶进行光合作用参数测定,测定时间为每天9:00~11:00 am 进行测量,光合测量的叶室条件设置为1 000 mmol·m-2·s-1光强,380±10 μmol·mol-1,温度为25±2 ℃,70%相对湿度。光合速率photosynthetic rate(Pn)、气孔导度 Stomatal conductance(Cond)、胞间 CO2浓度 Intercelular CO2concentration(Ci)、蒸腾速率 Transpiration rate(Tr)由光合仪直接导出,每个处理玉米幼苗叶片测量3 次[20]。
1.3.2 叶绿素及叶绿素荧光测定
采用OS-30p 叶绿素脉冲制荧光分析仪(美国Opti-Sciences 公司)对玉米幼苗的第二片展开叶进行叶绿素荧光特性测定。暗适应30 min 后读取初始荧光Fo,待Fo稳定后再照射饱和脉冲光,可得最大荧光Fm,光系统II(PSII)最大光化学量子效率[(Fv/Fm)=(Fm-Fo)][20]。
采用丙酮浸提比色法对玉米幼苗第二片展开叶进行测定,首先,称取新鲜叶片0.1 g,加入50 mL 提取液(80%丙酮和无水乙醇等体积混合),将提取液放置于避光处静止,待叶片完全褪色后,对提取液进行测定,测定其在470、646 nm 和 663 nm 下的OD值,并根据相关公式对玉米幼苗叶片的叶绿素含量进行计算[21]。
1.3.3 抗氧化酶活性测定
玉米叶片 MDA 含量,O2-产生速率,H2O2含量,可溶性蛋白含量,SOD、APX、GPX、GR、POD、CAT 活性测定参照文献[22],以上指标均测定同一处理下3株玉米植株叶片,即每个处理测定3 次重复,其中抗氧化酶活性以每毫克蛋白质所具有的酶活力单位数表示U·mg-1,测定时间为施用木霉菌后27 d。
1.4 数据分析
所有试验数据均采用SPSS 21.0 进行统计分析,Duncan 多重比较分析不同处理的影响差异,利用Microsoft Excel 2003 绘制表格。
2 结果与分析
2.1 盐碱胁迫下哈茨木霉和长枝木霉对玉米幼苗叶片光合作用参数的影响
盐碱胁迫(Con)显著降低了ZJ8 和JY417 两种玉米幼苗的光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Cond),同时使两品种玉米幼苗叶片胞间CO2浓度(Ci)显著升高,表明盐碱胁迫对ZJ8 和JY417 叶片的光合作用的限制效应可能以非气孔因素为主(表2)。施用不同木霉菌处理后,显著提高了ZJ8 和JY417的光合特性,其中,与盐碱胁迫(Con)处理相比,长枝木霉处理(TL)下ZJ8 和JY417 玉米幼苗叶片光合速率、蒸腾速率、气孔导度分别提高了74.11%、34.04%、42.70%和 53.47%、30.26%、30.35%,胞间 CO2浓度下降了43.69%和59.51%;哈茨木霉处理(TH)下分别提高了70.35%、30.54%、34.60%和47.86%、27.64%、24.34%,胞间CO2浓度下降了41.15%和56.91%。结果表明,不同木霉菌处理下,显著缓解了寒地盐碱土壤对玉米幼苗的胁迫效应,长枝木霉菌处理效果较好,且对‘ZJ8’缓解作用大于‘JY417’。
2.2 盐碱胁迫下哈茨木霉和长枝木霉对玉米幼苗叶片光合色素含量及叶绿素荧光参数的影响
光合色素含量高低对作物光合作用能力强弱具有非常大的影响。由表3(a and b)可知,盐碱胁迫(Con)显著降低 ZJ8 和 JY417 叶片 Chl a、Chl b、Chl a+b 和Car 含量。而施用不同木霉菌后,显著缓解了盐碱胁迫对 ZJ8 和 JY417 光合色素含量的影响(P<0.05),长枝木霉其中处理(TL)下ZJ8 和JY417 玉米幼苗叶片Chl a、Chlb、Chl a+b 和 Car 分别提高了 44.92%、82.26%、49.25%、41.84%和30.66%、73.32%、35.78%、39.45%;哈茨木霉处理(TH)下分别提高了38.30%、70.40%、42.02%、33.75%和23.67%、63.96%、29.50%、32.26%。
盐碱胁迫显著影响了ZJ8 和JY417 叶片叶绿素荧光参数(表3b),这表明,盐碱胁迫下对ZJ8 和JY417 叶片光合作用PSⅡ系统产生显著影响。施用哈茨木霉和长枝木霉后,ZJ8 和 JY417 叶片 Fv/F0、Fv/Fm均显著提高(P<0.05),长枝木霉处理(TL)下 ZJ8和 JY417 玉米幼苗叶片 Fv/F0、Fv/Fm分别提高了77.88%、14.68%和58.83%、12.41%;哈茨木霉处理(TH)下分别提高了72.56%、11.55%和54.11%、9.97%,以上结果说明盐碱胁迫下,不同木霉菌对玉米幼苗叶片光合作用PSII 反应中心的光化学活性具有显著缓解效果,从而促进玉米叶片的光合作用效率,长枝木霉处理效果较好,且对ZJ8 的效应更为显著。
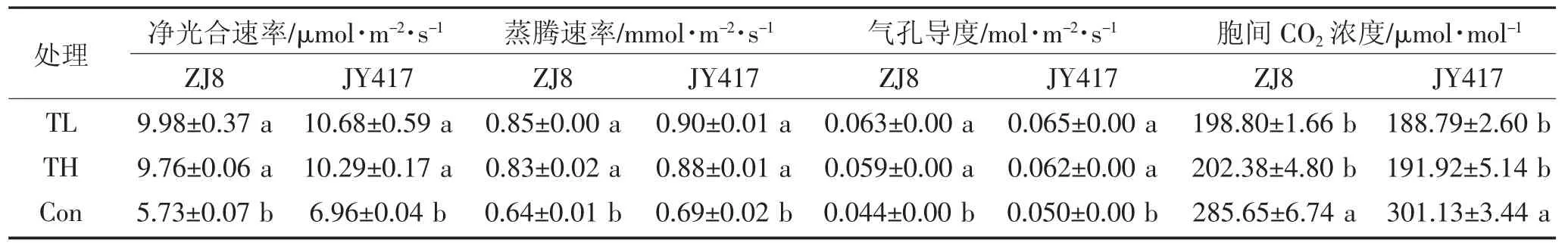
表2 不同木霉菌对盐碱土玉米幼苗叶片光合特性的影响Table 2 Influence of different Trichoderma on the photosynthetic parameters in the leaves of maize seedlings grown on saline-alkaline soil

表3a 不同木霉菌对盐碱土玉米幼苗叶片光合色素含量及叶绿素荧光参数的影响Table 3a Effect of different Trichoderma on photosynthetic pigments content and chlorophyll fluorescence parameter in maize seedlings leaves on saline-alkaline soil
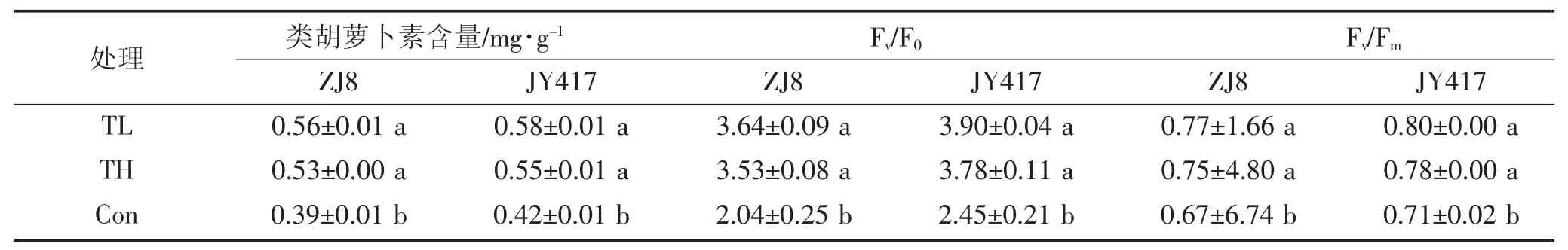
表3b 不同木霉菌对盐碱土玉米幼苗叶片光合色素含量及叶绿素荧光参数的影响Table 3b Effect of different Trichoderma on photosynthetic pigments content and chlorophyll fluorescence parameter in maize seedlings leaves on saline-alkaline soil
2.3 盐碱胁迫下哈茨木霉和长枝木霉对玉米幼苗叶片ROS 积累及膜脂过氧化的影响
盐碱胁迫(Con)下,ZJ8 和 JY417 叶片的 MDA 和H2O2含量及O2-产生速率均显著增加(P<0.05)。施用不同木霉菌后降低了各处理叶片中H2O2和O2-的积累(表4),缓解了盐碱胁迫诱导的膜质过氧化,从而使MDA 的积累量显著降低。与盐碱胁迫处理相比,长枝木霉(TL)处理下ZJ8 和JY417 玉米幼苗叶片H2O2、O2-、MDA 含量降低了 51.85%、46.87%、63.64%和49.29%、43.64%、57.98%;哈茨木霉(TH)处理下降低了 46.52%、39.24%、60.19%和 44.21%、34.80%、53.92%。研究表明土壤中施用不同木霉菌后能提高不同玉米对盐碱胁迫的抗性,长枝木霉处理效果较好,对ZJ8 缓解较为明显。

表4 不同木霉菌对盐碱土玉米幼苗叶片ROS 的积累及脂质过氧化的影响Table 4 Influence of different Trichoderma on the accumulation of ROS and on the oxidation parameters in the leaves of maize seedlings in saline-alkaline soil
2.4 盐碱胁迫下哈茨木霉和长枝木霉对玉米幼苗叶片渗透调节物质的影响
渗透调节物质有助于调节逆境条件下植株渗透压,防止细胞脱水。盐碱胁迫(Con)处理下,ZJ8 和JY417 两种玉米幼苗叶片中的脯氨酸和可溶性糖含量显著低于木霉菌处理(P<0.05)(表5)。施用不同木霉菌后ZJ8 和JY417 两种玉米幼苗叶片的渗透调节物质显著提高,与对照(Con)相比,长枝木霉(TL)处理下ZJ8 和JY417 玉米幼苗叶片脯氨酸和可溶性糖含量提高了17.50%、15.55%和40.10%、35.28%;哈茨木霉(TH) 处理下含量提高了14.75%、12.09%和37.86%、31.79%,且可溶性糖含量对缓解盐碱胁迫效果好于脯氨酸。

表5 不同木霉菌对盐碱土玉米幼苗叶片可溶性渗透调节物的影响Table 5 Influence of different Trichoderma on the content of osmolytes in the leaves of maize seedlings in saline-alkaline soil
2.5 盐碱胁迫下哈茨木霉和长枝木霉对玉米幼苗叶片抗氧化酶活性的影响
与木霉菌处理相比,盐碱胁迫(Con)显著降低了ZJ8 和 JY417 两种玉米幼苗叶片的 SOD、APX、GPX、GR 和 POD 活性(P<0.05),但 CAT 活性变化规律与其他抗氧化酶活性有所不同,盐碱胁迫(Con)下,ZJ8和JY417 两种玉米叶片CAT 酶活性较木霉菌处理显著提高。不同品种间酶活性的含量也表现出不同差异,JY417 酶活性均高于ZJ8。施用不同木霉菌后,ZJ8和 JY417 玉米叶片的 SOD、POD、APX、GPX、GR 的活性均高于盐碱胁迫处理,而CAT 活性则相反,长枝木霉(TL)处理下ZJ8 和JY417 玉米幼苗叶片SOD、POD、APX、GPX、GR 酶活性提高了 26.52%、23.49%、79.37%、65.01%、67.89%和24.58%、21.50%、73.73%、52.72%、55.71%,CAT 活性降低了11.46%和10.60%;哈茨木霉(TH) 处理下酶活性提高了 24.51%、21.29%、73.51%、58.89%、59.63%和22.72%、19.77%、70.71%、50.89%、51.43%,CAT 活性降低了 10.21%和8.15%,长枝木霉处理效果较好,且对ZJ8 缓解明显。
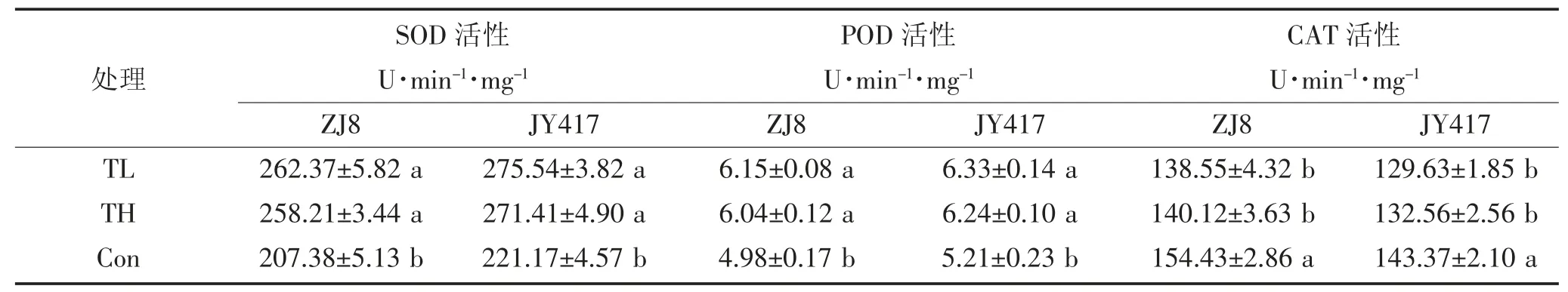
表6a 不同木霉菌对盐碱土玉米幼苗叶片抗氧化酶活性的影响Table 6a Influence of different Trichoderma on the antioxidant enzyme activities in the leaves of maize seedlings in saline-alkaline soil

表6b 不同木霉菌对盐碱土玉米叶片抗氧化酶活性的影响Table 6b Influence of different Trichoderma on the antioxidant enzyme activities in the leaves of maize seedlings in saline-alkaline soil
3 讨论
作物的光合作用对作物产量具有非常重要的作用,而盐碱土壤胁迫对作物的光合作用具有非常显著的抑制作用,从而影响作物的正常生长发育[23]。研究发现,ZJ8 和JY417 两种玉米幼苗叶片的光合速率、气孔导度和蒸腾速率在盐碱胁迫均显著降低,而叶片胞间CO2则显著升高(表2);而哈茨木霉和长枝木霉处理下,ZJ8 和JY417 两种玉米幼苗叶片的光合速率、气孔导度和蒸腾速显著升高,而叶片胞间CO2则显著降低。这说明,两种不同木霉菌对盐碱胁迫下ZJ8 和JY417 两种玉米幼苗叶片光合作用的提高主要通过缓解叶片非气孔限制因素来实现。
作物叶片的细胞膜透性对盐碱胁迫非常敏感,从而导致膜透性升高,抑制作物叶片的叶绿素合成,同时使叶片叶绿素降解速度加快,最终使作物的光合作用受到显著抑制[24]。研究中发现,ZJ8 和JY417两种玉米幼苗叶片的光合色素含量在盐碱胁迫下均显著下降(表3a and b),这表明盐碱胁迫下,叶片Chl 的降解加快,而叶绿素合成受到明显抑制,导致作物叶片叶绿体的光合作用降低;盐碱胁迫同时ZJ8和JY417 两种玉米幼苗叶片的细胞膜透性显著增加,导致细胞膜脂过氧化,是玉米发生叶片发黄枯萎的状态。而施用哈茨木霉和长枝木霉后,ZJ8和JY417两种玉米幼苗叶片的光合色素含量显著提高,同时缓解了盐碱胁迫导致的叶片细胞膜透性增加,降低了叶片膜脂过氧化,基于此,木霉菌通过缓解不同品种玉米叶片的膜脂过氧化程度,提高叶片光合色素含量,这与前人研究结果相似[25],研究结果同时发现木霉菌处理对ZJ8 的缓解作用较明显。
研究作物叶片荧光参数在环境胁迫下的变化规律可以有效的探析逆境胁迫对作物光合作用结构损害程度,同时可以帮助研究人员掌握作物对光能的利用效率[26]。研究结果表明,ZJ8 和JY417 两品种玉米幼苗叶片的Fv/F0、Fv/Fm在盐碱胁迫下显著下降(表3b),表明不同品种玉米幼苗光能利用率在盐碱胁迫下均被显著降低,最终使玉米的生长发育受到抑制。盐碱胁迫下,施用哈茨木霉和长枝木霉能够有效提高ZJ8 和JY417 两品种玉米幼苗叶片的Fv/F0、Fv/Fm,从而有效缓解盐碱胁迫对不同玉米叶片光合作用PSII 反应中心活性、光能利用效率产生的损伤,进而提高不同玉米品种的光合能力,对ZJ8 缓解作用更好,这一结果与研究中ZJ8 和JY417 两品种玉米幼苗叶片的光合参数的测定结果相吻合。
在盐碱胁迫下,通过不同木霉菌处理后,增强了玉米叶片的光合特性和光合色素含量,并诱导渗透调节物质可溶性糖,脯氨酸含量的变化。可溶性糖、脯氨酸可以有效促进渗透调节,从而导致脱水耐受性增强。通过降低渗透势来进行渗透调节,在细胞的水分保持和膨胀维持中起着关键作用,从而通过平衡溶质势来最大程度地降低盐碱胁迫的不利影响,有助于细胞生长。同时,不同木霉菌处理诱导了酶的变化,如 SOD、POD、APX、GPX 和 GR 酶活性的增加。以最大程度地降低盐碱胁迫下ROS 引起的氧化损伤,作为多种防御机制之一,酶促抗氧化剂系统在提高植物抗性中起着重要作用。在抗氧化酶水平的提高可保护作物组织免受盐碱胁迫下的膜氧化损伤,从而减轻盐碱胁迫毒害改善玉米植株的生长。SOD、POD 和CAT 是作物抗氧化防御的关键成分[27]。作为抑制ROS 产生的第一种酶,SOD 是参与盐碱胁迫耐受性的最有效的酶促抗氧化剂。基于这些机制的累积作用,盐碱胁迫下,施用哈茨木霉和长枝木霉能够有效缓降低胁迫对不同玉米品种幼苗造成的损伤。在促进不同品种玉米幼苗生长的同时,导致H2O2、O2-、MDA 含量的减少。简而言之,木霉菌处理可能通过增加ROS 清除,并在盐碱胁迫下维持渗透平衡和代谢稳态来提高玉米的耐盐碱性。
在生理水平上,木霉菌处理下缓解盐碱胁迫对玉米生长的影响也归因于作物中的ROS 水解(表4)。在正常条件下,包括超氧阴离子(O2-)、过氧化氢(H2O2)在内的ROS 在低浓度下可作为信号分子,但快速而过量的ROS 积累会在盐胁迫下损害植物。 ROS 会影响细胞膜的完整性,酶活性和植物的光合作用设备[27-28]。 ROS 引起的氧化胁迫是暴露于环境胁迫下的植物的主要损害之一[29]。为了保护免受氧化胁迫的损害,作物依靠非酶和酶系统清除过量的ROS。例如增加酶促和非酶促抗氧化剂以清除ROS,并提高植物对盐胁迫的耐受性[30]。因此,抗氧化防御系统通过调节这些酶以及在施用木霉菌后盐碱胁迫下的抗氧化剂水平,在提高耐受性和减少有害作用方面起着重要作用。
4 结论
不同木霉菌处理后,显著促进了两玉米杂交品种的生长,提高了玉米幼苗的耐盐碱性。研究结果发现哈茨木霉和长枝木霉处理均有效提高了盐碱胁迫下不同基因型玉米幼苗叶片光合特性及光合色素含量,增加了ROS 的清除能力,维持渗透平和和代谢稳态。结果表明,不同木霉菌处理能够有效缓解盐碱土壤胁迫对作物的危害,因此可以将木霉菌开发成为可持续农业的有效生物肥料。
猜你喜欢
作物学报(2022年8期)2022-05-29
当代水产(2022年1期)2022-04-26
金桥(2021年11期)2021-11-20
安徽林业科技(2021年6期)2021-03-05
东坡赤壁诗词(2018年2期)2018-05-10
科学Fans(2017年12期)2018-01-03
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
少儿科学周刊·少年版(2015年3期)2015-07-07
