
热休克蛋白27 通过调节氧化应激影响奶牛胎衣不下发生机制研究
2021-03-11 07:56张梦龙赵璧忱邢菲菲罗胜缤罗春海付世新
黑龙江八一农垦大学学报 2021年1期
张梦龙,赵璧忱,邢菲菲,罗胜缤,罗春海,付世新
(黑龙江八一农垦大学动物科技学院,大庆 163319)
胎衣不下(Retention of fetal membranes,RFM),是指胎儿胎盘在产犊后8 到12 h 之内没有排出,其降低了牛的生产力和繁殖力,对乳制品业造成重大经济损失[1-2]。已知一些易患RFM 的风险因素,但是由于其多因素病因,故统一的发病机制仍然难以寻找。分娩后胎儿胎膜的分离以及排出主要与牛胎盘的结构和免疫学变化有关[3],而已经研究证明类固醇激素,妊娠晚期和分娩时的前列腺素环境在一定程度上介导了这些变化[4]。近年来随着技术的发展,研究者们对于该疾病的研究着手于更为直观的影响因素上,例如影响胎盘分离的蛋白质[5],如 FAK[6]、MMPs[5]、VEGFA[7-8]等,影响子宫肌肉收缩的 Ca2+、STIM1 等。在实验室的前期研究中,发现在RFM 与NO-RFM 奶牛胎盘中存在一种潜在的重要差异蛋白—小热休克蛋白[9]。
热休克蛋白27(HSP27 或HSPB1)是一种应激蛋白,属于小分子热休克蛋白家族中的一员,当细胞处于能够改变蛋白折叠的条件下(例如热刺激)时[10],其表达水平会上调,目前HSP27 可能是研究最深入的小分子热激蛋白之一。HSP27 于1987 年至1988 年被首次纯化和鉴定,在这之后人们很快发现HSP27具有形成高分子量寡聚物并且能够磷酸化的能力。几年后,发现HSP27 是一种发挥作用不依赖于ATP的分子伴侣,其参与了蛋白质的折叠机制,在这一过程中的主要作用是捕获和储存由于应激诱导形成的错误折叠的多肽,以避免其聚集,并间接的促进其再折叠或错误蛋白的降解。Hsp27 也在细胞骨架组织的控制中扮演了重要角色,并以其抗凋亡和抗氧化特性而闻名。而由于胎盘的功能是为胎儿提供营养和保护,因此具有极高的代谢活性,在妊娠过程中,线粒体数量增加表明氧代谢可能会很高,但升高的氧代谢可能会导致产生大量的活性氧(ROS),其中包括超氧阴离子自由基(O2-)过氧化氢(H2O2)和极活泼的羟基(OH-)。ROS 能够破坏细胞的分子结构,例如核酸,脂质和蛋白质,故而可能会诱导细胞死亡和(或)触发凋亡途径。而目前已有研究表明,当奶牛发生RFM 时,胎儿胎盘与母体胎盘中过氧化氢酶(CAT)和超氧化物歧化酶(SOD)等与ROS 代谢相关的酶活性出现极显著差异[11-12],这证明奶牛的RFM 与ROS可能有极大的联系[13]。
而HSP27 的抗氧化特性也是广为人知的,该特性是由于其可产生针对氧化应激诱导物和病理环境的物质,进而对细胞起到保护作用[14]。研究发现HSP27 能够降低细胞内活性氧(ROS)和一氧化氮(NO)的水平。而早已经有研究发现谷胱甘肽(GSH)本身具有非常强大的抗氧化能力,同时有研究者发现HSP27 能够调节6-磷酸葡萄糖脱氢酶(G6PD)的活性[15],那么当奶牛发生胎衣不下时,差异表达的HSP27 在胎盘之间扮演一种怎样的角色?其与胎盘之间的ROS 有怎样的联系?HSP27 是否通过调节G6PD 进而影响GSH 的产生?故而实验对HSP27 对RFM 奶牛与正常奶牛胎盘,以及子宫内膜上皮细胞的氧化应激的调节进行了研究探讨。
1 材料与方法
1.1 试剂
Western 及IP 细胞裂解液、蛋白酶抑制剂混合物、BCA 蛋白浓度测定试剂盒、GSH 和GSSG 检测试剂盒、谷胱甘肽过氧化物酶检测试剂盒、活性氧检测试剂盒、G6PDH 活性检测试剂盒,SDS-PAGE 凝胶配制试剂盒,青霉素-链霉素溶液,均购买于碧云天生物技术有限公司;胰蛋白酶-EDTA 消化液,4×蛋白上样缓冲液均购自于上海索莱宝公司;RPMI 1640 液体培养基购自于Hyclone 公司;胎牛血清购自于四季青公司;Anti-HSP27(Abcam,鼠源,单抗)购自于美国Abcam 公司;Anti-Actin Beta(云克隆,鼠源,单抗)购自于中国武汉云克隆公司;Goat Anti-Mouse(纳川,山羊)购自于中国哈尔滨纳川生物公司。Si-RNA,siRNA-Mate 转染试剂均购自于苏州吉玛基因公司;17-AAG 购自于上海陶素生化公司。
1.2 样品采集
在黑龙省某现代化奶牛养殖场进行样品采集,选取一批品系,体况评分,产次,产奶量都相近的奶牛,并且在其分娩后对其进行消毒及局部麻醉后,手术采集胎儿胎盘,并且根据RFM 判断标准以及对奶牛进行跟踪以观察是否有其他疾病的发生,最终选取胎衣不下与正常奶牛各7 头,RFM 记为T 组,正常组记为C 组;在手术采集时应注意采集的组织大小适中,采集后立刻使用灭菌生理盐水冲洗血污,至于液氮中保存,之后迅速运回实验室至于-80 ℃保存。
1.3 Western Blot
研钵提前灭菌备用,液氮预冷,剪取适量组织放入其中,使用研杵将其研磨成干粉状,在此过程中务必保证研钵中始终有液氮,用药匙将干粉倒入EP 管中,加入裂解液提取组织总蛋白,然后使用BCA 测定盒检测蛋白浓度,并将其调至相同,加入蛋白上样缓冲液煮沸变性后备用,根据目的蛋白大小选择浓度合适的分离胶,按照说明书配制,根据蛋白浓度调整上样量,根据marker 标记切取凝胶,使用半干法转膜,之后使用5%脱脂奶粉封闭,结束后使用TBST 清洗,选择相对应的一抗孵育过夜,之后孵育二抗,每个步骤之间都需要TBST 清洗,最后使用凝胶成像系统照膜,Image Lab 软件分析灰度值。
1.4 G6PD、GSH-Px、GSH 和 GSSG 的检测
根据各个WST-8 试剂盒的说明书,分别检测胎衣不下与胎衣正常胎儿胎盘中G6PD 及GSH-Px 的活性,GSH 和 GSSG 含量。
1.5 细胞培养及处理
奶牛原代子宫内膜上皮细胞来自于实验室,子宫内膜上皮细胞在含10%血清,1%青链霉素的培养基中,外界环境为5% CO2以及37 ℃条件下培养。
实验室应用H2O2法已经成功建立奶牛子宫内膜上皮氧化应激模型,并且也通过预实验筛选了HSP27 过表达剂17-AAG[16]的适宜用量;故而进行了以下处理:使用17-AAG 对HSP27 进行过表达,再使用过氧化氢溶液处理细胞;使用未经任何处理,并且处于氧化应激状态下的细胞作为对照,使用上述方法检测细胞中HSP27 的表达,G6PD、GSH-Px 的活性以及ROS、GSH 和GSSG 的含量变化。以上试验每次实验设置3 个重复,并且每个试验重复3 次。
1.6 统计分析
结果使用平均值±SEM 表示,使用SPSS 20.0 软件进行分析,使用单因素ANOVA 方法对数据进行分析,“*”表示差异显著(P<0.05),“**”表示差异极显著(P<0.01)。
2 结果
2.1 组织总蛋白的提取与浓度测定结果
提取组织蛋白后,应用BCA 法测定蛋白浓度,并绘制蛋白标准曲线,其中R2=0.993 4≈1,表明得出标准曲线准确性高。然后根据标准曲线计算出各个样品的浓度如表1。

表1 组织样品总蛋白浓度及吸光度值Table 1 The concentration and OD of protein in the groups
2.2 组织蛋白Western blot 结果
Western blot 实验结果见图1,使用分析软件可知HSP27 在胎衣不下奶牛胎儿胎盘中显著的上调(P<0.05),并且 HSP27 蛋白大量活化。
2.3 组织中G6PD、GSH-Px 活性以及GSH 和GSSG的含量检测
由表2 可见,当奶牛发生胎衣不下时,胎盘中G6PD 的活性出现显著性下降(P<0.05),而 GSH-Px活性则出现相反的结果,在胎衣不下胎盘中呈现显著性上调(P<0.05)。

图 1 HSP27 与 p-HSP27 的 Western Blot 结果Fig.1 The result of HSP27 and p-HSP27 Western blotting
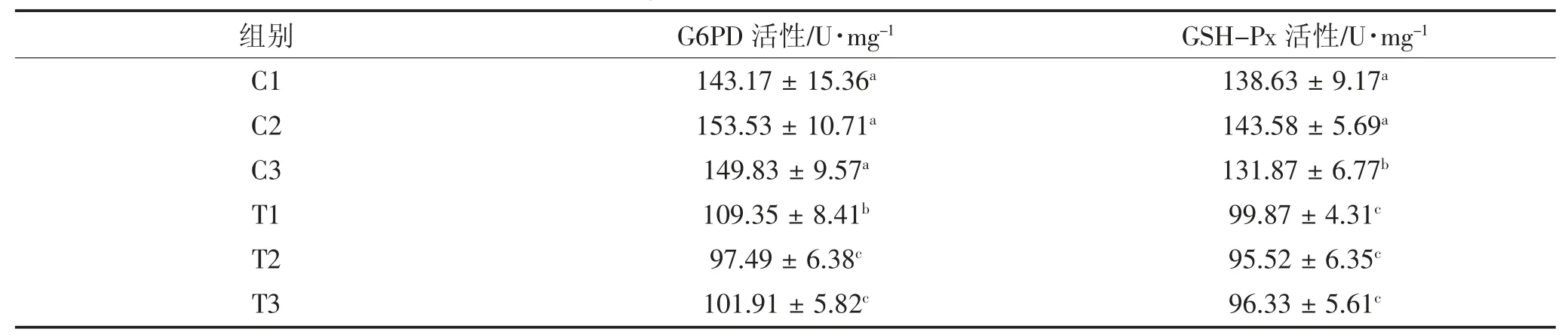
表2 组织中G6PD 与GSH-Px 的活性Table 2 Activity of G6PD and GSH-Px in the caruncles
由表3 可知,奶牛在正常分娩时胎盘中GSH 含量较高,GSSG 含量较低,而当奶牛发生胎衣不下时GSH 含量与正常相比呈现显著性降低(P<0.05),GSSG 含量显著性增高(P<0.05)。

表3 胎盘中GSSG 与GSH 的含量Table 3 Content of GSSG,GSH in the caruncles
2.4 过表达HSP27 对牛子宫内膜上皮细胞氧化应激的影响
2.4.1 过表达HSP27 后,其在氧化应激下的表达量
由图2 可以得知,实验成功的对HSP27 进行了过表达。
2.4.2 过表达HSP27 对牛子宫内膜上皮细胞ROS 含量的影响
HSP27 的过表达显著减少了由于H2O2刺激诱导产生的 ROS(P<0.01)。
2.4.3 过表达HSP27 对牛子宫内膜上皮细胞G6PD、GSH-Px 活性的影响
由表4 可知,对HSP27 的过表达显著增强了由于 H2O2刺激而降低的 G6PD 活性(P<0.01),GSH-Px活性的活性变化呈现与G6PD 相似的结果(P<0.01)。
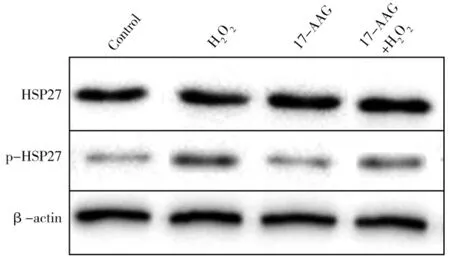
图2 过表达HSP27 对子宫内膜上皮细胞HSP27 表达的影响Fig.2 Effect of overexpression of HSP27 on HSP27 abundance in UCE

图3 过表达HSP27 对牛子宫内膜上皮细胞ROS 含量的影响Fig.3 Effect of overexpression of HSP27 on the content of ROS in UCE

表4 过表达HSP27 对子宫内膜上皮细胞G6PD 及GSH-Px 活性的影响Table 4 Effect of overexpression of HSP27 on the activity of SOD and GSH-Px in UCE
2.4.4 过表达和沉默HSP27 对牛子宫内膜上皮细胞GSH 和 GSSG 的含量检测
由表5 可知,对HSP27 的过表达显著降低了细胞中由于 H2O2刺激而增加的 GSSG 含量(P<0.01),GSH活性的含量变化呈现于GSSG 相反的结果(P<0.01)。

表5 过表达HSP27 对子宫内膜上皮细胞GSSG 及GSH 含量的影响Table 5 Effect of overexpression of HSP27 on the content of GSSG and GSH in UCE
3 讨论
奶牛胎衣不下本身是一种多因素疾病,目前的研究涉及了更为微观的层次,例如胎盘在分离以及排出过程中一些分子的变化,大致概括为免疫反应、炎症反应,细胞外基质的重塑以及细胞凋亡等。而实验验证并研究了在RFM 奶牛及子宫内膜上皮细胞中HSP27 对其氧化应激的影响,从HSP27 通过调节G6PD 进而影响GSH 的合成的途径对氧化应激的调节从而探讨其对奶牛分娩后胎衣排出的影响。
HSP27 在生物体内发挥作用主要通过磷酸化和特异的寡聚化两种途径;研究发现当生物体处于氧化应激状态时[17],HSP27 迅速形成较大分子的寡聚化结构(>400 kDa),过一段时间后HSP27 会逐渐转化为较小的寡聚化结构(<200 kDa),而寡聚化的HSP27 主要发挥其“分子伴侣”的功能[18],在细胞处于氧化应激状态时,修复发生错误折叠的蛋白,将其维持在折叠状态,并且可以使受损不严重的多肽链重新折叠;保护细胞骨架以及降低DNA 损伤[19]。而细胞处于氧化应激时,大分子量寡聚化的HSP27 甚至可能会增加ATP 依赖性伴侣分子(Hsp70)的效率,而实验中发现,当奶牛发生胎衣不下时,HSP27 的表达量上升,但是大部分以磷酸化形式存在;其寡聚化分子量显著降低,所以HSP27 可能通过上述功能在一定程度上影响胎衣的排出。
之前的研究发现寡聚化的可以刺激6-磷酸葡萄糖脱氢酶(G6PD)的活性[20],而 G6PD 是戊糖磷酸途径的第一个酶,在细胞中催化能量代谢,该酶也是限速酶,而这一途径是红细胞中还原型辅酶Ⅱ(NADPH)的唯一来源,而实验发现,HSP27 寡聚化的减少,降低了G6PD 的活性,可能导致了NADPH 的减少,使得红细胞中H2O2的堆积,最终导致氧自由基的产生,破坏红细胞,导致胎盘局部缺氧坏死,使得胎盘无法正常分离;而其G6PD 不仅仅参与戊糖磷酸途径,也参与糖酵解途径,两种途径中都产生ATP,由于HSP27 降低了其活性,导致ATP 产生减少,使得奶牛在分娩时由于子宫收缩无力易发生难产[21],而在产后更难以正常排出胎衣,故而极大的增加了RFM 的发生概率。
戊糖磷酸途径的生物学功能远不止于此[22],在该途径中生成的NADPH 会与氧化型谷胱甘肽(Glutathione Disulfide,GSSG)经由谷胱甘肽还原酶(Glutathione Reductase,GS)生成还原型谷胱甘肽(Glutathione,GSH),GSH 是一种含半胱氨酸的三肽(γ-Glu-Cys-Gly),在真核细胞中广为存在,浓度大约在0.5~10 mM,而当细胞处于氧化应激状态时,硒依赖性谷胱甘肽过氧化物酶(GSH-Px)通过利用GSH 还原有机过氧化物以及H2O2,维持细胞正常的氧化水平,而GSH 则被氧化变为GSSG,GSH 的还原能力足以对细胞的氧化损伤提供一定的保护作用;而研究发现当奶牛发生RFM 时,GSH-Px 的活性出现显著性的变化,实验中降低的HSP27 寡聚化蛋白使得其未能发挥“分子伴侣”的功能,可能导致了几种酶活性的变化;由于G6PD 活性的降低,使得能量及NADPH 生成减少,最终导致GSH 生成不足,而使得奶牛处于严重的氧化应激状态下,极大的增加胎衣无法正常排出可能性。
而当机体处于氧化状态时,还易生成不饱和脂肪酸-亚油酸(Linolic acid)和花生四烯酸(ARA),它们是部分重要激素,如前列腺素合成的前体;GSHPx 还通过催化前列腺素G2(PGG2)向前列腺素H2(PGH2)转化进而直接参与花生四烯酸的代谢[23]。而机体大量的ROS 可能导致PGG / H 合酶不可逆的失活,并且由于细胞调用了大量的GSH-Px 减少ROS的产生[24],从而可能影响前列腺素的浓度,已经研究发现前列腺素浓度的失常易导致RFM。
综上所述,HSP27 对于氧化应激状态下奶牛胎衣的排出与分离是一个多过程的影响:首先,HSP27本身对于氧化应激状态具有一定的调节功能,其次HSP27 在氧化应激状态下,对G6PD 以及GS 活性的调节,进而影响GSH 的生成,而这一过程中的酶以及最终产物GSH 都能对ROS 发挥一定的调节作用,进而协同影响胎衣的排出。
猜你喜欢
现代农村科技(2022年7期)2022-11-18
吉林畜牧兽医(2022年7期)2022-11-16
临床肝胆病杂志(2022年10期)2022-10-19
现代仪器与医疗(2021年6期)2022-01-18
农民致富之友(2019年16期)2019-07-01
风湿病与关节炎(2016年12期)2017-01-14
科学与管理(2016年5期)2016-12-01
中国中药杂志(2016年20期)2016-11-19
家庭百事通·健康一点通(2016年9期)2016-09-21
大众健康(2015年2期)2015-09-10
