
松毛虫赤眼蜂两性生殖品系和孤雌产雌品系在不同品种柞蚕卵中的寄生和生长发育表现
2021-03-10 02:23:54周金成
昆虫学报 2021年1期
周金成,何 玥,赵 倩,董 辉
(沈阳农业大学植物保护学院,沈阳 110866)
以柞蚕Antheraeapernyi[鳞翅目(Lepidoptera):天蚕蛾科(Saturniidae)]为卵替代寄主繁育的松毛虫赤眼蜂Trichogrammadendrolimi[膜翅目(Hymenoptera):赤眼蜂科(Trichogrammatidae)]已被广泛应用于防治多种鳞翅目害虫(张俊杰等,2015;Zhouetal.,2019a)。相比于其他寄主昆虫的卵,柞蚕卵作为替代寄主具有营养丰富、繁蜂效率高、价格低廉、易于获得、便于贮藏及运输等诸多优势(Zhangetal.,2018)。我国有着悠久的柞蚕养殖历史,经过长期选育及遗传改良,已具有丰富的柞蚕种质资源(李玉忠,1994;Zhouetal.,2020)。不同柞蚕品种的卵内营养物质及性状具有差异性,将对松毛虫赤眼蜂的寄生及发育存在潜在影响(石生林等,2016)。因此,筛选繁育赤眼蜂的最佳柞蚕卵品种是赤眼蜂大卵繁育技术的最上游环节,对改善繁蜂效率和质量至关重要。
寄主质量是影响寄生蜂寄生及生长发育的关键因子(Godfray,1994)。首先,当寄主质量较低时,用于子代蜂发育的寄主营养匮乏,导致子代蜂个体变小,繁殖力下降,发育畸形甚至死亡,而当寄主营养较丰富时,子代蜂则倾向于将剩余营养资源分配于卵巢和脂肪体,育出的子代蜂体型增大,繁殖力增强。此外,脂肪及糖原等营养物质储量增加也有利于提高赤眼蜂抗逆性,提高其田间生存能力,还可以延长子代蜂低温贮藏时间(王生等,2011)。其次,寄主质量会对赤眼蜂的寄生选择和寄生率产生影响。窝卵数理论(clutch size theory)认为,寄生蜂可通过判断寄主营养质量,在一次寄生时产下适当的子代数量。当寄主营养丰富时,寄生蜂会倾向于分配较多的子代,而对低质量的寄主,寄生蜂会选择仅分配少量的子代甚至不产卵(Lack,1947;Charnov and Skinner,1985;Zhouetal.,2020)。因此,替代寄主质量将对赤眼蜂规模化繁育效率及成本产生重要影响(Wajnbergetal.,2016)。
赤眼蜂对柞蚕卵的寄生及其子代生长发育不仅受柞蚕卵质量影响,同时也受不同蜂种或蜂品系的营养需求规律及发育特点的影响。例如,胞内共生菌Wolbachia可通过诱导赤眼蜂单倍体未受精卵第一次有丝分裂失败,导致胚胎染色体加倍,进而育出二倍体雌性后代,即发生孤雌产雌现象(Miura and Tagami,2004)。由于雌蜂无需交配就能产下几乎100%具有寄生功能的子代雌蜂,因而孤雌产雌品系的赤眼蜂具有繁蜂效率高、野外定殖力强、控害潜能高等优势(Stouthamer,1993;Liuetal.,2019)。然而,Wolbachia的侵染将对子代蜂的发育及代谢均产生影响,进而影响其对寄主营养的需求规律(Russell and Stouthamer,2011)。因此,筛选适于繁育孤雌产雌品系松毛虫赤眼蜂的最佳柞蚕卵品种将有利于改善其规模化繁育技术。
为筛选适于繁育两性生殖品系和孤雌产雌品系松毛虫赤眼蜂的最佳柞蚕卵品种,并初步揭示两种品系子代蜂的营养需求特点,本研究统计并分析了两种品系雌蜂寄生6种品种柞蚕卵的寄生率、子代蜂窝蜂数(单窝羽化子代蜂数)、子代蜂个体大小和子代性比;通过检测不同品种柞蚕卵的蛋白质、甘油三酯、总糖含量和单卵湿重等指标,明确不同品种柞蚕卵的营养质量;利用主成分分析方法,探究柞蚕卵营养指标与两种品系赤眼蜂生物学指标的相关性。研究结果将为改善松毛虫赤眼蜂规模化繁育技术提供新的思路,并为筛选适于赤眼蜂繁育的优良柞蚕品种卵提供依据与数据支撑。
1 材料与方法
1.1 供试昆虫
本试验所用的两性生殖品系和感染Wolbachia的孤雌产雌品系松毛虫赤眼蜂均来源于同一遗传背景的自然种群。孤雌产雌品系松毛虫赤眼蜂为Wolbachia经显微注射转染至两性生殖的雌蜂后诱导获得。
6个品种的柞蚕卵由沈阳市新丰生物科技服务有限公司提供,分别为抗大(KD)、大四(DS)、高新(GX)、988(NEE)、青大(QD)和特大(TD)。供试柞蚕卵均为目前工厂化繁育赤眼蜂普遍应用的未受精雌蛾的剖腹卵,由羽化后1日龄的柞蚕雌蛾腹部卵巢中获得,经去离子水漂洗,筛除绿卵、病卵后晾干备用。
本研究所需的供试昆虫种群均在25±1℃,相对湿度70%±5%,光周期16L∶8D条件下连代饲养。
1.2 赤眼蜂生物学指标的测定
为避免雌蜂初次寄生不同品种柞蚕卵的不适应性对试验结果的影响,本试验预先分别以6种品种柞蚕卵为寄主连续饲养两种品系赤眼蜂3代。取饲养第4代1日龄雌蜂,每40头雌蜂分别接入至预先放有30粒各品种柞蚕卵的指形管(直径2 cm,长10 cm)寄生24 h后移出雌蜂。将寄生后的柞蚕卵单粒单管分装,每日观察子代蜂羽化情况,未羽化的柞蚕卵剖查其卵内是否存在遗留子代蜂。各赤眼蜂品系寄生每品种柞蚕卵各设3次重复,记录各品种柞蚕卵寄生率。每一处理组合下90粒卵寄生后分别记录其窝蜂数(单窝羽化子代蜂数)和雌性比,每卵各随机选取3头羽化雌蜂,测量其后足胫节长度指示其个体大小。
1.3 柞蚕卵质量指标的测定
1.3.1单卵湿重测定:取羽化1日龄的柞蚕雌蛾,去头后解剖雌蛾腹部卵巢,经去离子水漂洗后筛除绿卵和病卵后自然晾干,以万分之一天平称取柞蚕卵湿重,每一柞蚕卵品种各设30个重复。
1.3.2蛋白质、总糖和甘油三酯含量测定:挑选不同品种的新鲜柞蚕卵粒各10粒,用灭菌后的解剖针刺破卵壳后,用移液枪吸取卵液收集于2 mL离心管内,在-80℃条件下贮藏备用。每处理以每10粒柞蚕卵为一个生物学重复,蛋白质、总糖和甘油含量的测定每一柞蚕品种设5个生物学重复。
柞蚕卵液可溶性蛋白含量采用考马斯亮蓝G-250染色法(王生等,2011;吴晓艺,2012)。取0.1 mL卵液于2 mL离心管,以去离子水定容至2 mL,充分振荡匀浆后以5 000 r/min离心20 min,取上清,管内沉淀再补入去离子水定容至2 mL,再次振荡匀浆、离心,合并上清。取1 mL上清液在595 nm波长下读取吸光值。预先以牛血清蛋白作为标准蛋白溶液梯度稀释后测定吸光值并绘制标准曲线。以标准曲线计算样品可溶性蛋白含量。
柞蚕卵液总糖含量的测定采用蒽酮比色法(王生等,2011;吴晓艺,2012),并略作改进。取0.1 mL卵液于10 mL离心管,加入1 mL去离子水,充分振荡匀浆后,加入2 mL 10%三氯乙酸振荡摇匀,以10 000 r/min离心10 min,取上清,管内沉淀再补入2 mL 10%三氯乙酸,再次振荡匀浆、离心,合并上清。取上清0.5 mL,以去离子水定容至1 mL,冷水浴中加蒽酮0.3 mL和浓硫酸3 mL,加热至沸腾约10 min,冷却后静置20 min后在620 nm波长下读取吸光值。同时预先以葡萄糖作为标准溶液梯度稀释后测定吸光度并绘制标准曲线。以标准曲线计算样品总糖含量。
甘油三酯含量使用南京建成甘油三酯(TG)试剂盒测定(王生等,2011;吴晓艺,2012)。取0.1 mL卵液于2 mL离心管,以去离子水定容至2 mL,充分振荡匀浆后以2 500 r/min离心10 min,取上清,管内沉淀再补入去离子水定容至2 mL,再次振荡匀浆、离心,合并上清。以2.26 mmol/L的标准甘油溶液为校准溶液,分别将去离子水、上清液和校准溶液加入试剂盒预混酶制剂37℃下孵育10 min后,以酶标仪(Versa Max MD,美国)测定吸光值。
1.4 数据分析
本研究采用广义线性模型(generalized linear model)分析了柞蚕品种和蜂品系对松毛虫赤眼蜂雌蜂寄生率和子代蜂个体大小的影响和柞蚕品种对两性品系松毛虫赤眼蜂子代蜂性比的影响。由于柞蚕卵质量指标数据均不满足模型正态性假定,采用KW秩和检验(Kruskal-Wallis rank sum test)分析了柞蚕品种和蜂品系对松毛虫赤眼蜂子代蜂窝蜂数的影响以及不同柞蚕品种卵内蛋白质含量、总糖含量、甘油三酯含量及单卵湿重的差异(Hall,1992)。同一试验因素下不同处理水平间的多重比较采用z检验估计差异显著性(显著性水平α=0.05)。采用主成分分析(principal component analysis)提取主成分,明确柞蚕卵质量指标与松毛虫赤眼蜂生物学指标的共相关性。提取特征值大于1的主成分,相关系数绝对值大于0.5视为存在共相关性。本研究所有数据均使用R统计软件进行分析(R Developmental Core Team,2018)。
2 结果
2.1 柞蚕品种和赤眼蜂品系对松毛虫赤眼蜂生物学指标的影响
柞蚕品种(F5,24=1.47,P=0.24)、蜂品系(F1,24=0.48,P=0.50)及两因素的交互作用(F1,24=0.89,P=0.50)对雌蜂对柞蚕卵的寄生率均无显著影响(图1)。

图1 柞蚕品种和蜂品系对松毛虫赤眼蜂雌蜂寄生率的影响Fig.1 Parasitism rate of female wasps of Trichogramma dendrolimi as influenced by Antheraea pernyi strain and wasp line图中数据为平均值±标准误;柱上不同小写和大写字母分别表示两性生殖品系和孤雌产雌品系雌蜂在不同品种柞蚕卵上的生物学参数差异显著(P<0.05,z测验);ns,星号和双星号分别表示两性生殖品系和孤雌产雌品系雌蜂寄生同一品种柞蚕卵时的生物学参数无显著差异(P>0.05),差异显著(P<0.05)和差异极显著(P<0.01)(z测验)。Data in the figure are mean±SE.Different lowercase and uppercase letters above bars indicate significant differences (P<0.05,z-test) in the biological parameters of bisexual and thelytokous female wasps,respectively,on eggs of different strains of A.pernyi.ns,single asterisk and double asterisk above bars indicate no significant difference (P>0.05),significant difference (P<0.05),and extremely significant difference (P<0.01)(z-test),respectively,in the biological parameter between bisexual and thelytokous female wasps on the eggs of the same strain of A.pernyi.图2和3同The same for Figs.2 and 3.
子代蜂个体大小受蜂品系和柞蚕卵品种间的交互作用显著影响(F5,171=5.68,P<0.001)。在两性品系子代蜂中,NEE品种柞蚕卵育出的松毛虫赤眼蜂子代蜂个体最大(271.10±15.80 μm),极显著大于DS(194.43±7.41 μm),GX(202.24±5.94 μm)和TD(191.57±5.87 μm)品种柞蚕卵育出的松毛虫赤眼蜂子代蜂的个体(DS:z=4.92,P<0.001;GX:z=4.60,P<0.001;TD:z=4.52,P<0.001),但与KD(221.23±5.15 μm)和QD(249.69±20.51 μm)品种柞蚕卵育出的松毛虫赤眼蜂子代蜂个体相比无显著差异(KD:z=2.74,P=0.068;QD:z=0.56,P=0.99)。在孤雌产雌品系子代蜂中,各品种柞蚕卵育出的松毛虫赤眼蜂子代蜂的个体大小均无显著差异(P>0.05),DS,GX,KD,NEE,QD和TD品种柞蚕卵育出的松毛虫赤眼蜂子代蜂个体大小分别为202.71±5.55,187.99±5.88,209.45±6.30,207.30±6.86,216.72和191.12±7.55 μm。NEE和QD品种柞蚕卵育出的两性品系松毛虫赤眼蜂子代蜂个体大小显著大于其育出的孤雌产雌品系子代蜂个体大小(NEE:z=4.27,P=0.001;QD:z=5.66,P<0.001),但DS,GX,KD和TD品种柞蚕卵育出的孤雌产雌品系松毛虫赤眼蜂子代蜂个体大小与其育出的两性品系松毛虫赤眼蜂子代蜂相比均无显著差异(DS:z=0.67,P=0.50;GX:z=1.20,P=0.23;KD:z=0.87,P=0.38;z=0.026,P<0.98)(图2)。

图2 柞蚕品种和松毛虫赤眼蜂品系对子代蜂个体大小的影响Fig.2 Body size of offspring wasps as influenced by Antheraea pernyi strain and Trichogramma dendrolimi line
Kruskal-Wallis检验结果表明,在孤雌产雌品系子代蜂中,KD品种柞蚕卵育出的松毛虫赤眼蜂子代蜂窝蜂数平均值最高(119.94±12.08头/卵),显著高于NEE(77.26±6.71头/卵)和QD(97.77±6.99头/卵)品种柞蚕卵育出的松毛虫赤眼蜂子代蜂的(NEE:P<0.001;QD:P=0.037),但与DS(107.63±6.90头/卵)、GX(105.71±9.18头/卵)和TD(114.50±5.45头/卵)品种育出的松毛虫赤眼蜂子代蜂窝蜂数相比无显著差异(DS:P=0.20;GX:P=0.38;TD:P=0.44)。在两性品系子代蜂中,不同品种柞蚕卵育出的松毛虫赤眼蜂子代蜂窝蜂数间均无显著差异(P>0.05),DS,GX,KD,NEE,QD和TD品种柞蚕卵育出的松毛虫赤眼蜂子代蜂窝蜂数分别为79.00±7.45,85.72±6.22,96.71±5.27,77.00±7.48,85.93±9.01和57.13±15.99头/卵。DS,GX和TD品种柞蚕卵育出的孤雌产雌品系松毛虫赤眼蜂子代蜂窝蜂数均显著高于其同品种柞蚕卵育出的两性品系松毛虫赤眼蜂子代蜂窝蜂数(DS:P=0.0074;GX:P=0.019;TD:P=0.021),但KD,NEE和QD品种柞蚕卵育出的两种品系松毛虫赤眼蜂子代蜂窝蜂数均无显著差异(KD:P=0.11;NEE:P=0.96;QD:P=0.16)(图3)。
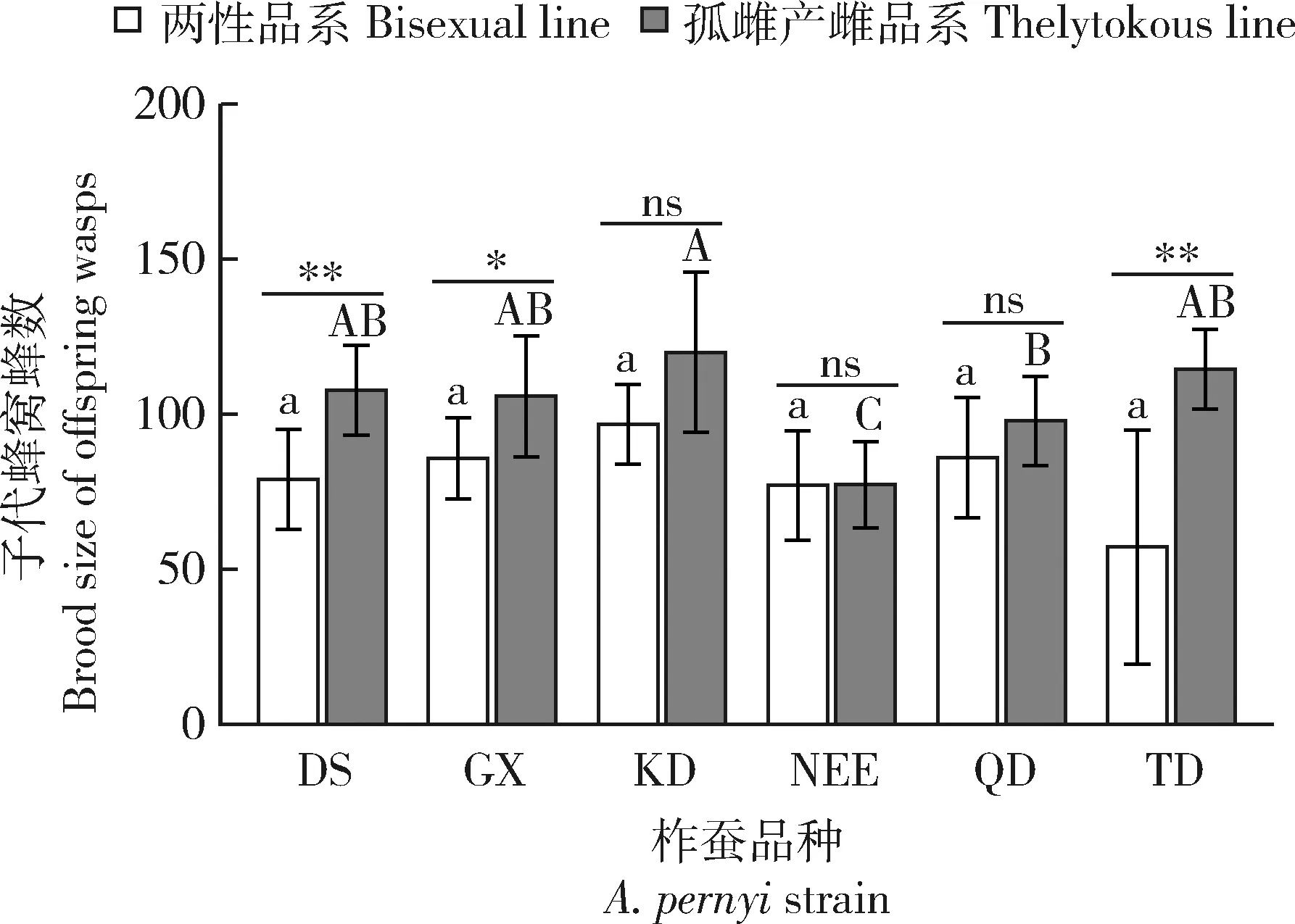
图3 柞蚕品种和蜂品系对松毛虫赤眼蜂子代蜂窝蜂数的影响Fig.3 Brood size of offspring wasps as influenced by Antheraea pernyi strain and Trichogramma dendrolimi line
柞蚕品种对两性品系松毛虫赤眼蜂子代蜂性比无显著影响(F5,64=0.088,P=0.99)(图4)。
2.2 不同品种柞蚕卵湿重及卵内各营养成分含量
Kruskal-Wallis检验结果表明,柞蚕品种对卵内蛋白质含量存在显著影响(χ2=15.44,df=5,P=0.0087)。TD品种柞蚕卵的蛋白质含量平均值最高(86.70±1.17 mg/mL),显著高于QD(79.48±0.90 mg/mL),DS(78.91±0.52 mg/mL),NEE(67.60±0.43 mg/mL)和KD(57.38±0.73 mg/mL)品种柞蚕卵的(QD:P=0.0020;DS:P<0.001;NEE:P<0.001;KD:P<0.001),但与GX品种(81.80±1.69 mg/mL)柞蚕卵的相比无显著差异(P=0.081)(图5:A)。

图4 不同品种柞蚕卵育出的两性品系松毛虫赤眼蜂子代蜂雌性比Fig.4 Proportion of female offspring wasps of bisexual line of Trichogramma dendrolim developed on eggs of different strains of Antheraea pernyi图中数据为平均值±标准误;柱上相同小写字母表示不同品种柞蚕卵内两性品系子代蜂雌性比差异不显著(P>0.05,z测验)。Data in the figure are mean±SE.The same lowercase letters above bars indicate no significant differences (P>0.05,z-test) in the proportions of female offspring wasps of bisexual line developed on eggs of different strains of A.pernyi.
柞蚕品种对卵内总糖含量存在显著影响(χ2=16.14,df=5,P=0.0064)。KD品种柞蚕卵的总糖含量显著高于其他品种柞蚕卵的(P<0.05)。NEE品种柞蚕卵总糖含量显著高于除KD品种外的其余品种柞蚕卵的(P<0.05)。QD品种柞蚕卵的总糖含量最低,显著低于其余品种柞蚕卵的(P<0.05)(图5:B)。
柞蚕品种对卵内甘油三酯含量存在显著影响(χ2=15.55,df=5,P=0.0083)。QD品种柞蚕卵的甘油三酯含量(70.26±1.77 mmol/L)显著高于其他品种柞蚕卵的(P<0.05)。KD品种柞蚕卵的甘油三酯含量最低(42.33±0.60 mmol/L),显著低于其余品种柞蚕卵的(P<0.05)(图5:C)。

图5 不同品种柞蚕卵内的蛋白质(A)、总糖(B)和甘油三酯(C)含量Fig.5 Contents of protein (A),total carbohydrate (B),and triglyceride (C) in eggs of different strains of Antheraea pernyi图中数据为平均值±标准差;柱上不同小写字母表示不同品种柞蚕卵内各营养物质含量差异显著(P<0.05,Kruskal-Wallis检验)。Data in the figure are mean±SE.Different lowercase letters above bars indicate significant differences in the nutrient contents in eggs of different strains of A.pernyi (P<0.05,Kruskal-Wallis test).
柞蚕品种对柞蚕卵单卵湿重存在显著影响(F5,174=42.06,P<0.001)。DS品种柞蚕卵的单卵湿重显著高于其余品种柞蚕卵(P<0.05)。GX和QD品种柞蚕卵单卵湿重较低,均显著低于其余4品种柞蚕卵(P<0.05)(图6)。
2.3 柞蚕卵质量指标与赤眼蜂生物学指标的主成分分析
提取的柞蚕卵质量指标与两性品系赤眼蜂发育指标的两个主成分变量共解释了73.40%的变异。其中,主成分1特征值为3.13,解释了44.74%的变异,主成分2特征值为2.01,解释了28.65%的变异。主成分1分别与两性品系松毛虫赤眼蜂寄生率(相关系数:0.85)、柞蚕卵蛋白质含量(0.94)和甘油三酯含量(0.65)呈正相关,但与柞蚕卵总糖含量(-0.95)呈负相关。主成分2分别与两性品系松毛虫赤眼蜂子代蜂个体大小(0.90)和柞蚕卵甘油三酯含量(0.69)呈正相关,但与柞蚕卵单卵湿重(-0.74)呈负相关(表1)。
提取的柞蚕卵质量指标与孤雌产雌品系松毛虫赤眼蜂生物学指标的两个主成分变量共解释了69.93%的变异。其中,主成分1特征值为2.66,解释了37.93%的变异;主成分2特征值为2.24,解释了32.00%的变异。主成分1分别与孤雌产雌品系松毛虫赤眼蜂寄生率(0.74)和柞蚕卵甘油三酯含量(0.95)呈正相关,但与松毛虫赤眼蜂子代蜂窝蜂数(-0.61)、柞蚕卵总糖含量(-0.54)和单卵湿重(-0.54)呈负相关。主成分2与孤雌产雌品系松毛虫赤眼蜂子代蜂个体大小(0.73)和柞蚕卵总糖含量(0.71)呈正相关,与柞蚕卵蛋白质含量呈负相关(-0.92)(表2和图7)。

图6 不同品种柞蚕卵的单卵湿重Fig.6 Wet weight per egg of different strains of Antheraea pernyi图中数值为平均值±标准误;柱上不同小写字母表示不同品种柞蚕卵的单卵湿重差异显著(P<0.05,Kruskal-Wallis检验)。Data in the figure are mean±SE.Different lowercase letters above bars indicate significant differences in the wet weight per egg between different A.pernyi strains (P<0.05,Kruskal-Wallis test).
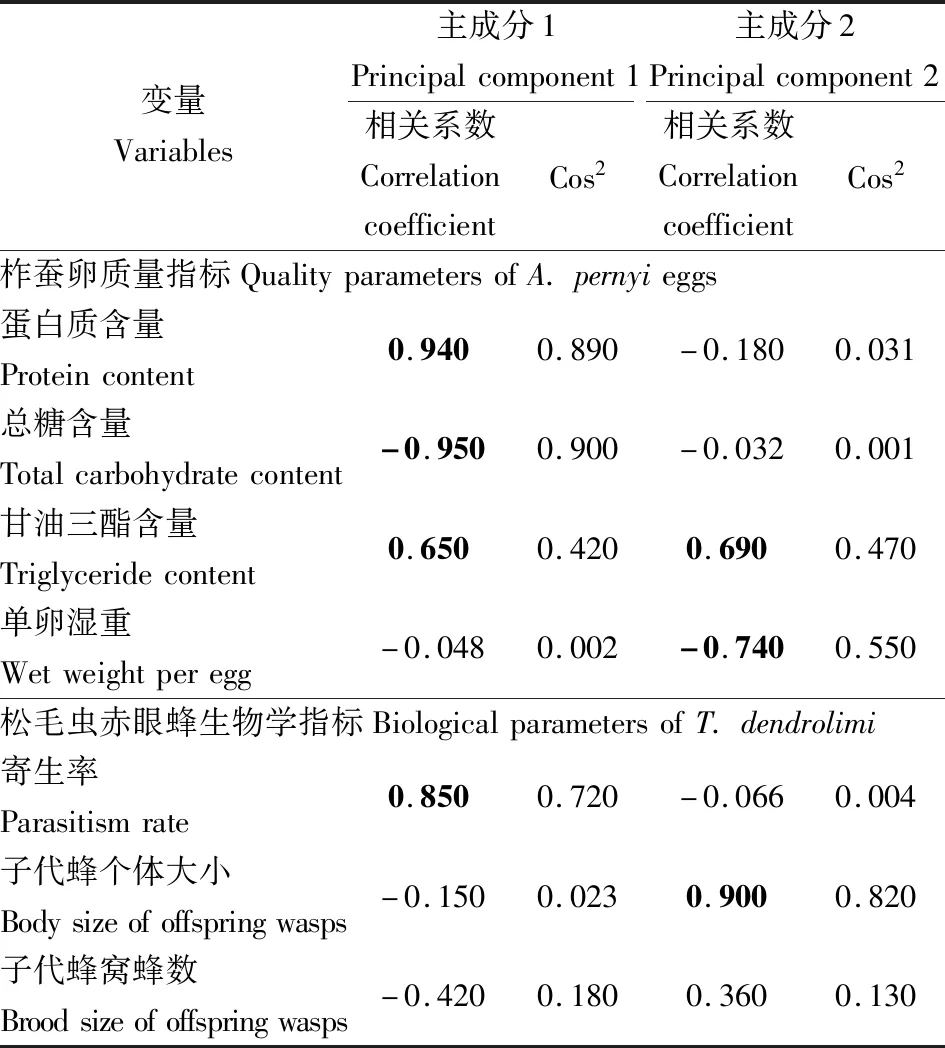
表1 两性品系松毛虫赤眼蜂生物学指标和柞蚕卵质量指标的主成分分析Table 1 Principal component analysis of the biological parameters of bisexual Trichogramma dendrolimi and the quality parameters of Antheraea pernyi eggs
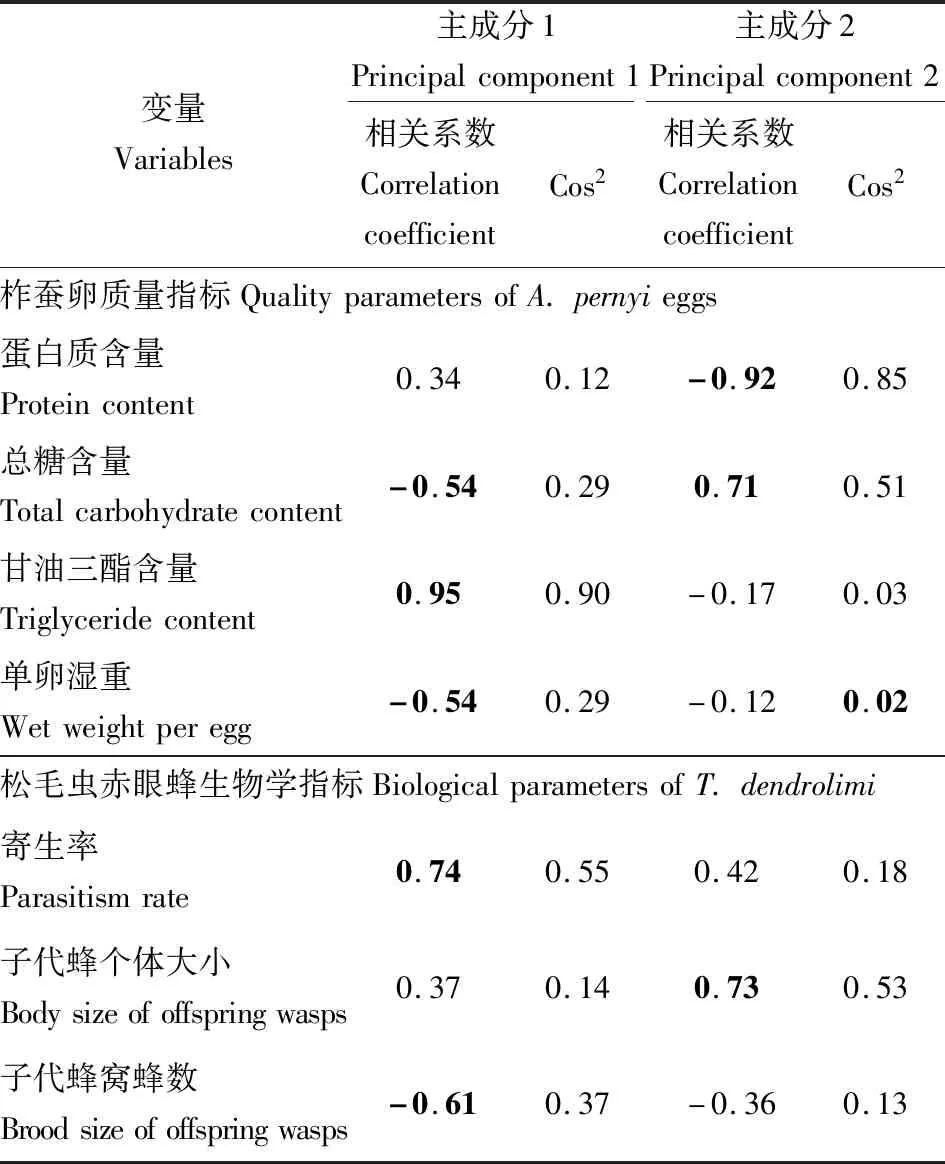
表2 孤雌产雌品系松毛虫赤眼蜂生物学指标和柞蚕卵质量指标的主成分分析Table 2 Principal component analysis of the biological parameters of thelytokous Trichogramma dendrolimi and the quality parameters of Antheraea pernyi eggs
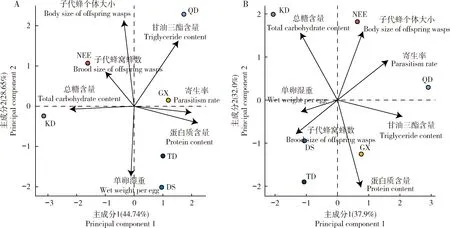
图7 两性品系(A)和孤雌产雌品系(B)松毛虫赤眼蜂生物学指标分别和柞蚕卵质量指标的双标图Fig.7 Bi-plots of the quality parameters of Antheraea pernyi eggs and biological parameters of bisexual (A) and thelytokous (B) lines of Trichogramma dendrolimi
3 讨论与结论
个体大小是反映生物适合度的重要指示变量,而窝蜂数则反映了单卵可育蜂数,与寄生率一同反映出规模化繁育过程中的繁蜂效率。一般而言,在保障发育的基础上,子代蜂会将剩余资源分配于卵巢和脂肪体,并增大其体型。因此,体型较大的子代蜂一般贮存有丰富的脂肪体(为羽化后的潜在寿命提供能量)并具有较高的繁殖力(更多数量的卵巢管和卵子)。当寄主可利用营养资源有限时,子代蜂为保证其完成发育,仅消耗少量寄主资源,导致个体变小或畸形,甚至有时会选择放弃翅的形成,产生残翅或缺翅现象(王生等,2011;张俊杰,2015;Zhouetal.,2019b)。因此,子代蜂适合度是寄主营养和雌蜂寄生选择相互平衡的结果。
本研究发现NEE和QD品种柞蚕卵繁育的两性品系松毛虫赤眼蜂子代蜂个体较大,但其窝蜂数不受柞蚕品种影响,而孤雌产雌品系雌蜂在DS,GX,KD和TD品种柞蚕卵上育出的子代蜂窝蜂数较高(图2和3)。相关性分析表明,两种品系松毛虫赤眼蜂的子代蜂个体大小和寄生率均与柞蚕卵甘油三酯含量呈正相关关系(表2;图7)。我们推测不同柞蚕卵对两种品系松毛虫赤眼蜂子代蜂发育的影响受子代蜂营养需求规律和柞蚕卵营养成分的共同影响。寄主卵内的甘油三酯是脂肪类物质的主要贮存形式,是子代蜂胚胎发育的重要营养来源,对其卵巢发育及适合度存在重要影响。通过摄取脂类物质,赤眼蜂将寄主卵内的营养储存于体内,促进其体型增大,增强其适合度与抗逆性。例如滞育条件下的赤眼蜂体型显著增大,体内甘油三酯含量持续增加(王生等,2011;张俊杰,2015)。浅黄恩蚜小蜂Encarsiasophia寄生后的烟粉虱Bemisiatabaci体内脂质含量被持续消耗(李敏,2013)。本研究结果初步揭示了柞蚕卵内脂质含量是影响松毛虫赤眼蜂的重要因子,在赤眼蜂的规模化繁育过程中,可针对性地选择脂质含量较丰富的柞蚕卵品种。此外,在实际生产过程中,农户及柞蚕生产厂家对柞蚕品种的市场认可度不同,且不同柞蚕品种的繁育具有地域性特点。因此,不同品种的柞蚕卵是否易获取也是赤眼蜂繁育过程中需要考虑的重要因素。本研究检验的6个柞蚕品种中,988和抗大品种柞蚕应用相对较广(李玉忠,1994;朱有敏等,2012),可考虑分别作为规模化繁育两性品系和孤雌产雌品系松毛虫赤眼蜂的推荐品种。此外,本研究仅对6种柞蚕品种进行了评价,而我国现有约160个柞蚕品种,其中各地区实用品种约30余种(李玉忠,1994;朱有敏等,2012),因此未来仍有必要进一步发掘适于繁育松毛虫赤眼蜂的柞蚕品种。
研究结果还发现柞蚕卵单卵湿重与孤雌产雌品系松毛虫赤眼蜂雌蜂的寄生率均存在显著负相关关系,而与两性品系松毛虫赤眼蜂雌蜂的寄生率无显著相关性(表1和2;图7)。一般而言,寄生蜂总是倾向于寄生较大的寄主来获取更充足的营养,而本研究结果却与此规律相悖。我们推断这种现象可能由于较大的柞蚕卵往往卵壳硬度和厚度较大,使部分个体较小的雌蜂产卵器可能无法刺透卵壳,即使雌蜂能够刺透卵壳将子代产于卵内,子代蜂也可能无法成功咬破卵壳,顺利羽化。例如玉米螟赤眼蜂T.ostriniae因其产卵器较短,无法刺穿柞蚕卵壳,通过采用物理或化学手段将卵壳打磨或软化后,雌蜂方可实现对其寄生(夏邦颖和王敏慧,1979)。在本研究中,柞蚕卵繁育的孤雌产雌品系松毛虫赤眼蜂子代蜂窝蜂数高于其育出的两性品系赤眼蜂子代蜂的窝蜂数。有研究发现,Wolbachia感染的孤雌产雌品系松毛虫赤眼蜂雌蜂表现出更高频率的过寄生行为,即在已被寄生的寄主卵表现多次产卵的行为(Liuetal.,2018;Zhouetal.,2019b)。Wolbachia则可能通过诱导高频率的过寄生行为实现其在同一寄主内子代蜂个体间的传播,这可能是Wolbachia促进自身传播的潜在策略(Zhouetal.,2019b)。对宿主赤眼蜂而言,过高频率的过寄生行为将导致柞蚕卵内窝蜂数增加,个体间营养竞争加剧,存在个体适合度下降的风险。
本研究还发现孤雌产雌品系松毛虫赤眼蜂子代蜂个体大小与柞蚕卵蛋白质含量呈负相关,与柞蚕卵总糖含量呈正相关,而柞蚕卵蛋白质和总糖含量与两性品系松毛虫赤眼蜂子代蜂个体大小无明显相关关系(表2;图7)。这说明两种品系松毛虫赤眼蜂的营养需求规律可能不同。有研究发现,Wolbachia感染的孤雌产雌生殖的蚬蝶赤眼蜂T.kaykai和狭氏赤眼蜂T.deion的子代卵裂期及胚胎发育速度相对于两性品系子代明显延缓,胚胎期延缓约12 h(Tagamietal.,2001)。这种子代蜂生长发育的不同步可能使子代蜂存在不同的营养需求规律。例如,宿主黑腹果蝇Drosophilamelanogaster的线粒体浓度随Wolbachia感染滴度的增加而上升(Henry and Newton,2018)。另一研究则发现,Wolbachia感染促进了ATP合成相关基因的表达上调和氧胁迫反应(Groteetal.,2017)。这些现象暗示了Wolbachia感染的宿主具有较强的能量代谢活动,而糖类是生物体代谢活动可直接利用的能源物质。因此,Wolbachia可能使宿主子代蜂对糖类营养物质的需求增加。
本研究评价了不同品种柞蚕卵对两种品系松毛虫赤眼蜂子代适合度的影响,并以主成分分析在计算过程中进行了降维处理,结果指出了柞蚕卵内不同营养物质组成及卵重与松毛虫赤眼蜂适合度指标的潜在相关性,但可能会掩盖不同品种柞蚕卵的其他潜在影响。因此,仍有必要围绕寄主卵与赤眼蜂的营养互作关系开展更深入的研究,以证明两者间的因果关系。本研究一方面为筛选工厂化繁育松毛虫赤眼蜂的优良柞蚕卵品种提供了参考,另一方面则通过建立柞蚕卵营养指标与松毛虫赤眼蜂子代蜂适合度指标的相关性关系,为进一步筛选用于繁育松毛虫赤眼蜂的柞蚕卵品种提供了依据。通过发掘和利用我国柞蚕种质资源以改善赤眼蜂规模化繁育技术方法。
猜你喜欢
少年博览·小学低年级(2023年9期)2023-10-09 09:51:46
环境昆虫学报(2023年2期)2023-05-27 14:00:56
陕西农业科学(2022年6期)2022-08-10 02:00:30
中国森林病虫(2022年2期)2022-05-21 01:36:40
小星星·阅读100分(低年级)(2021年4期)2021-06-15 03:22:48
数学小灵通·3-4年级(2020年9期)2020-10-27 03:26:08
作文评点报·小学三、四年级(2020年13期)2020-04-20 11:19:13
浙江大学学报(农业与生命科学版)(2019年4期)2019-09-19 03:11:58
防护林科技(2018年2期)2018-03-09 07:54:52
现代农业(2015年5期)2015-02-28 18:40:49
