
5种赤眼蜂品系对米蛾卵和梨小食心虫卵的选择偏好研究
2022-08-10 02:00:30殷诗杰李先伟相会明刁红亮马瑞燕
陕西农业科学 2022年6期
殷诗杰, 李先伟, 相会明, 刁红亮, 马瑞燕
(山西农业大学 植物保护学院,山西 太谷 030801)
梨小食心虫Grapholitamolesta(Busck) 属鳞翅目 (Lepidoptera) 卷叶蛾科 (Tortricidae),简称“梨小”,是世界性的主要蛀果类害虫之一[1]。梨小主要危害核果类和部分仁果类果树,严重影响果实的品质和产量[2]。由于梨小主要以幼虫为害,且幼虫钻蛀取食果树的嫩梢和果实,极具有隐蔽性,常用的化学防治方法无法有效的杀灭幼虫,且容易产生抗药性和环境问题[3~5]。因此,在该虫卵期进行防治逐渐受到人们的关注。
赤眼蜂Trichogrammaspp. 是重要的卵寄生性天敌昆虫,寄主范围十分广泛。雌蜂产卵于寄主卵内,幼虫取食寄主卵液完成发育,并导致寄主死亡。在鳞翅目害虫的产卵高峰期通过“淹没式”释放赤眼蜂可以有效防治害虫的为害,特别是对于取食位置隐蔽的钻蛀性害虫而言,具有非常好的效果[6~7]。然而,每公顷田地释放赤眼蜂的数量往往需要达到数十万级才能达到良好的防治效果,因此直到人工繁育取得成功后,才出现较大规模的田间试验与示范[8]。赤眼蜂从卵到成虫之前都在寄主卵中生长发育,因此,选择合适的寄主卵成为了人工繁育赤眼蜂的关键[9]。我国经过长期的研究实践,已经成功实现用祚蚕Antheraeapernyi卵、米蛾Corcyracephalonica卵作为繁育寄主大量繁殖赤眼蜂[10~11]。目前,人工繁育的松毛虫赤眼蜂Trichogrammadendrolimi、玉米螟赤眼蜂Trichogrammaostriniae等已应用于玉米螟Ostriniafurnacalis、棉铃虫Helicoverpaarmigera、烟青虫Helicoverpaassulta等农林害虫的生物防治[9, 12]。
在生产应用中,赤眼蜂对目标害虫的偏好和寄生能力等会受到繁育寄主种类的影响[13~16]。张帆等 (2005) 研究显示,松毛虫赤眼蜂、螟黄赤眼蜂Trichogrammachilonis在柞蚕卵上培育3代后,对柞蚕卵的寄生率显著高于亚洲玉米螟卵的寄生率。Kaiser等报道赤眼蜂在非偏好寄主上饲养后会增强该种寄主对其的吸引力[18]。宋静等的研究表明羽化自二化螟Chilosuppressalis(Walker) 卵的稻螟赤眼蜂对二化螟卵表现出了寄主选择偏好,而来自米蛾卵的稻螟赤眼蜂对米蛾卵与二化螟卵均未表现出选择偏好[19]。
因此,为了更好地将赤眼蜂应用于田间梨小的防治,本实验研究了已商品化生产的松毛虫赤眼蜂和玉米螟赤眼蜂的不同品系,以及本地优势种暗黑赤眼蜂对梨小卵的选择趋性。研究结果为合理利用赤眼蜂防治梨小提供了一定理论参考。
1 材料与方法
1.1 材料
松毛虫赤眼蜂和玉米螟赤眼蜂的米蛾卵品系来自沈阳农业大学害虫生物防治研究室,在本实验室内分别用米蛾卵和梨小卵进行继代饲养10代以上,从而构建实验所用的松毛虫赤眼蜂和玉米螟赤眼蜂的米蛾卵品系和梨小卵品系。米蛾和梨小来源于山西农业大学生物安全与生物防治基地长期人工饲养的实验室种群,室内饲养方法参照已有的报道[20-21]。
暗黑赤眼蜂Trichogrammapintoi采自山西省晋中市太谷区周边桃园内被寄生的梨小卵,室内采用梨小卵进行继代饲养10代以上,以构建本地优势赤眼蜂种暗黑赤眼蜂的梨小卵品系,并用于本实验。共有五种赤眼蜂品系:松毛虫赤眼蜂米蛾卵品系(TD-CC)、 松毛虫赤眼蜂梨小卵品系(TD-GM)、玉米螟赤眼蜂米蛾卵品系(TO-CC)、玉米螟赤眼蜂梨小卵品系(TO-GM)、暗黑赤眼蜂梨小卵品系(TP-GM)。
1.2 方法
使用Y型嗅觉仪进行赤眼蜂的行为测定。Y型嗅觉仪主臂长10 cm,侧臂长10 cm,侧壁间夹角为45°。主臂和侧臂内部直径均为0.5 cm。寄主卵使用24 h以内产下的新鲜卵,赤眼蜂使用羽化后24 h以内的雌蜂。梨小卵产在1 cm×0.5 cm的硫酸纸上,同时把产在纱布上的米蛾卵扫下,并粘在相同大小的硫酸纸上。每一赤眼蜂品系均设置三个处理:①在“Y”型嗅觉仪两臂分别放置放有10粒米蛾卵的硫酸纸和干净硫酸纸;②在“Y”型嗅觉仪测臂分别放置放有10粒梨小卵的硫酸纸和干净硫酸纸;③分别将放置有10粒梨小卵的硫酸纸和放有10粒米蛾卵的硫酸纸放在“Y”型嗅觉仪两臂。使气流由两臂流向主臂,空气流速为200 mL/min。生测时,通过指形管将赤眼蜂逐头引入嗅觉仪的主臂内,观察记录10 min 内接入赤眼蜂的行为反应,并记录作出选择时所用的时间。判断标准为:当赤眼蜂爬至超过某侧臂5 cm 处,并持续1 min以上,记为此蜂对该侧臂内的物质做出了选择,否则记为未作选择。试验在25.2℃的室内进行,每测试5头赤眼蜂调换Y型嗅觉仪方位,每测试10头赤眼蜂更换新的寄主卵,每一实验处理重复不少于60次。
1.3 数据统计
所有试验数据采用SPSS25.0软件进行统计分析。使用2检验比较差异显著性。
2 结果与分析
2.1 五种赤眼蜂品系对米蛾卵的选择趋性

注:ns和*表示处理间无差异和差异显著
五种赤眼蜂品系对米蛾卵的行为反应测定结果见图1。松毛虫赤眼蜂米蛾卵品系对米蛾卵的选择数量低于选择对照的数量,但未达到显著差异水平 (χ2= 1.032;df= 1;P> 0.05);在其余四种赤眼蜂品系中,赤眼蜂对米蛾卵的选择量均高于对照,但仅在松毛虫赤眼蜂梨小卵品系中此种选择差异达到了显著性差异 (χ2= 7.172;df= 1;P< 0.05)。
2.2 五种赤眼蜂品系对梨小卵的选择趋性

注:ns表示处理间无显著差异
松毛虫赤眼蜂米蛾卵品系 (χ2= 0.300;df=1;P>0.05) 和梨小卵品系 (χ2= 0.248;df=1;P>0.05),以及玉米螟赤眼蜂梨小卵品系 (χ2= 2.036;df=1;P>0.05) 对梨小卵的选择数量高于选择空白对照的数量;玉米螟赤眼蜂米蛾卵品系 (χ2= 0.539;df=1;P>0.05) 和暗黑赤眼蜂梨小卵品系 (χ2= 1.200;df=1;P>0.05) 对梨小卵的选择数量低于选择对照的量,但在此五个处理中选择趋性的差异均未达到显著性水平(图2)。
2.3 五种赤眼蜂品系对两种繁育寄主的选择趋性
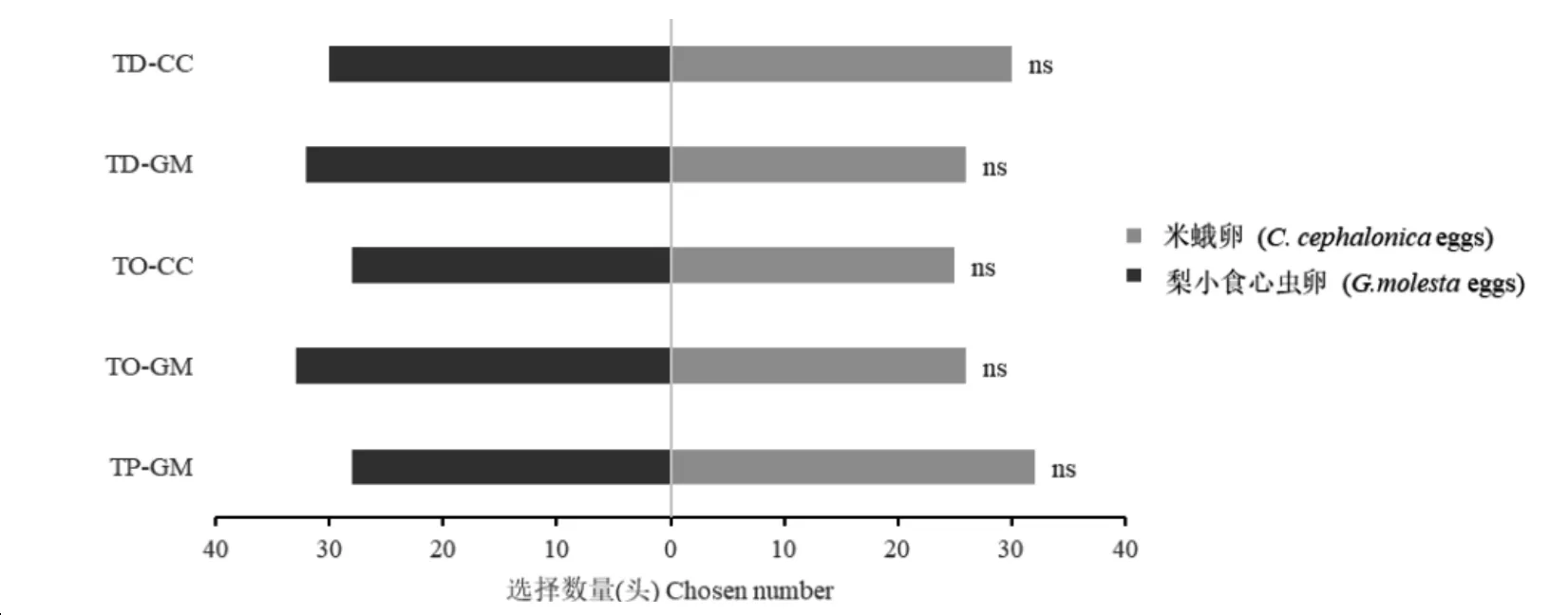
注:ns表示处理间无显著差异
五种赤眼蜂品系对米蛾卵及梨小卵的行为选择结果见图3。松毛虫赤眼蜂米蛾卵品系(χ2=0;df= 1;P> 0.05) 对米蛾卵和梨小卵的选择数量相同;松毛虫赤眼蜂梨小卵品系 (χ2= 0.776;df=1;P>0.05)、玉米螟赤眼蜂米蛾卵品系 (χ2= 0.304;df= 1;P>0.05) 和梨小卵品系 (χ2= 1.502;df= 1;P>0.05) 对梨小卵的选择量高于对米蛾卵的选择量;暗黑赤眼蜂梨小卵品系 (χ2= 0.533;df=1;P>0.05) 对梨小卵的选择数量低于选择米蛾卵的量。然而此五个处理中选择趋性的差异均未达到显著性水平。
2.4 五种赤眼蜂品系对米蛾卵的选择时间
五种赤眼蜂品系对米蛾卵的选择时间见图4。暗黑赤眼蜂梨小卵品系选择时间显著短于除玉米螟赤眼蜂梨小卵品系以外的三种赤眼蜂品系,而松毛虫赤眼蜂梨小卵品系对米蛾卵的选择时间最长,需170.9s (F = 3.665;df= 4;P< 0.05) 。
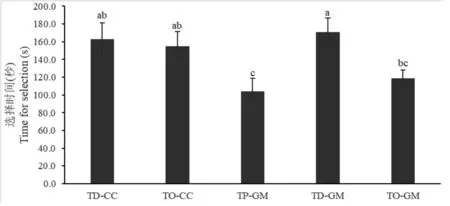
图4 不同赤眼蜂品系对米蛾卵的选择时间
2.5 五种赤眼蜂品系对梨小卵的选择时间
松毛虫赤眼蜂米蛾卵品系对梨小卵的选择时间最短,仅需77.2 s,而暗黑赤眼蜂梨小卵品系选择时间则显著长于两种松毛虫赤眼蜂品系及玉米螟梨小卵品系(F = 5.230;df= 4;P< 0.05) (图5)。

图5 不同赤眼蜂品系对梨小卵的选择时间
2.6 五种赤眼蜂品系对两种繁育寄主的选择时间
五种赤眼蜂品系同时面对两种繁育寄主时的选择时间见图6。玉米螟赤眼蜂米蛾卵品系选择时间显著长于其余四种赤眼蜂品系,需176 s(F=6.722;df= 4;P< 0.05)。其余四种赤眼蜂品系的选择时间差异不显著。

图6 不同赤眼蜂品系对米蛾卵和梨小卵的选择时间
3 结论与讨论
我们的研究表明除松毛虫赤眼蜂梨小品系选择米蛾的数量显著高于对照外,其余五种赤眼蜂品系对供试的两种寄主卵均未表现出明显的选择偏好。玉米螟赤眼蜂米蛾卵品系在三种处理中选择时间均较长。其余四种赤眼蜂在仅面对来自繁育寄主的气味时,作出选择的时间相对长于来自另一繁育寄主的赤眼蜂。
已有报道的寄主选择理论认为,赤眼蜂在羽化时需要咬破寄主卵,从而会“学习”到繁育寄主的气味,并在之后的寄主选择过程中表现出对该寄主的嗜好[22]。但本试验所测试的五种赤眼蜂品系却未观察到这种对繁育寄主的偏好。已有的研究表明,单寄主型寄生蜂可能更容易识别来自寄主的特异性线索,而多寄主型寄生蜂则倾向于利用一些更常见的线索来搜索潜在寄主[25~28]。如一种多寄主型寄生蜂缺肘反颚茧蜂Aphaeretaminuta(Nees) 并不通过识别来自寄主的化学物质搜索寄主幼虫,而是攻击在腐烂物质中遇到的几乎所有寄主[29]。因此我们推测本实验供试赤眼蜂对繁育寄主未表现出偏好的可能原因是,所试赤眼蜂均为多寄主型寄生性天敌。
研究结果表明,本地采集的暗黑赤眼蜂在所有三个试验处理中均表现出了最低的不选择量,即有着对气味信息物质的最高反应率,并且选择米蛾卵的数量要高于选择梨小卵或对照的数量。鉴于在利用天敌昆虫防治害虫时,有目的的使用相似生境中的天敌种类或品系,特别是需要注重本地天敌品系的筛选培养,更容易提高生物防治的成功率[30]。因此,我们认为这种本地采集到的暗黑赤眼蜂这一优势蜂种,有着使用米蛾卵进行人工培养,并应用于本地梨小生物防治的潜能。
在本试验中,五种赤眼蜂品系对梨小卵均未表现出明显的偏好或避忌行为反应。因此仅根据这一指标,此五种赤眼蜂品系尚不能确定可用于梨小田间防治的最优种,仍需要后期对寄生率、后代数量等指标做进一步的比较。
猜你喜欢
少年博览·小学低年级(2023年9期)2023-10-09 09:51:46
中国森林病虫(2022年2期)2022-05-21 01:36:40
今日农业(2021年14期)2021-11-25 23:57:29
小星星·阅读100分(低年级)(2021年4期)2021-06-15 03:22:48
数学小灵通·3-4年级(2020年9期)2020-10-27 03:26:08
今日农业(2020年14期)2020-08-14 01:10:20
作文评点报·小学三、四年级(2020年13期)2020-04-20 11:19:13
防护林科技(2018年2期)2018-03-09 07:54:52
现代农业(2016年5期)2016-02-28 18:42:38
现代农业(2016年5期)2016-02-28 18:42:34
