
楚雄腮扁叶蜂聚集产卵引诱物质初探
2019-09-19 03:11:58闫争亮马惠芬毛云玲刘凌胡光辉
浙江大学学报(农业与生命科学版) 2019年4期
闫争亮,马惠芬,毛云玲,刘凌,胡光辉
(云南省林业和草原科学院森林保护研究所,昆明 650201)
楚雄腮扁叶蜂(Cephalcia chuxiongicaXiao)隶属于膜翅目(Hymenoptera)扁叶蜂科(Pamphiliidae)腮扁叶蜂属(Cephalcia),仅在中国云南省有分布。1984年,肖刚柔首次报道了该物种,它是一种有较大经济重要性的害虫[1]。楚雄腮扁叶蜂主要危害云南松(Pinus yunnanensisFranch.)、华山松(Pinus armandiiFranch.)、滇油杉(Keteleeria evelynianaMast.)等重要的经济林木。该蜂以幼虫取食松树叶片,造成树木畸形、生长缓慢,为害严重时导致寄主死亡。该蜂2年发生1代,从6月下旬开始,成虫羽化,然后成虫发生期持续至10月上旬。其幼虫取食针叶时,吐丝形成灰白色的幕巢,且为害持续时间长达5个月之久,然后以老熟幼虫或预蛹在地表下10~20 cm处筑土室越冬越夏,长达17个月,直至第3年化蛹、羽化[2]。由于楚雄腮扁叶蜂独特的生物生态学特性,因此,对其防治较为困难。
我们前期的研究[3]表明:楚雄腮扁叶蜂已交尾雌蜂偏好在云南松、华山松上产卵,极少选择在滇油杉上产卵,但绝不会在云南拟单性木兰(Parakmeria yunnanensis)上产卵;而在其喜食寄主云南松、华山松针叶上喷施非寄主植物的树叶粗提物可抑制其产卵。室内试验表明:云南松松针粗提物、带卵松针粗提物、产卵雌蜂均对楚雄腮扁叶蜂已交尾雌蜂具有明显的引诱作用,且带卵松针粗提物的引诱作用明显大于不带卵的松针粗提物;此外,松针粗提物虽对楚雄腮扁叶蜂雄蜂具有一定的引诱作用,但与带卵松针粗提物的引诱作用间没有显著差异[4]。而且我们还观察到,雌蜂产卵于松树针叶以后的一段时间内,可诱集更多的同种雌蜂在同一个松梢上聚集产卵[3]。这些发现说明,研究利用信息化学物质对楚雄腮扁叶蜂进行防控的技术,是一个合理的思路。
以松叶蜂科(Diprionidae)为代表的广腰亚目(Symphyta)林木害虫的信息化学物质研究,主要聚焦在其雌蜂产生的性信息素上[5]。但由于性诱剂只对雄蜂起作用,而不能有效控制大量产卵的雌蜂,所以性信息素的应用对控制林间害虫虫口密度的效果不大。对广腰亚目林木害虫寄主挥发物的研究,则主要集中于害虫对适宜和不适宜寄主的辨别上。如欧洲新松叶蜂(Neodiprion sertifer)[6]和欧洲赤松叶锋(Diprion pini)[7-8]可以通过松树的挥发性物质的化学指纹图谱来区分不同的寄主;为害木质部的树蜂科(Siricidae)云杉树蜂(Sirex noctilio)的产卵雌蜂依据不同松树种类挥发物的化学指纹图谱决定其产卵偏好[9]。扁叶蜂科(Pamphiliidae)的松阿扁叶蜂(Acantholyda posticalis)可根据不同树种的气味区分适宜与非适宜寄主[10-11]。有研究发现:松叶蜂科的欧洲赤松叶蜂(D.pini)幼虫取食时口器分泌物可驱逐同种产卵雌蜂[12];欧洲新松叶蜂(Neodiprion sertifer)的卵淋洗液不会对其雌蜂产卵产生驱避作用[13],但新松叶蜂属的Neodiprion fulviceps产卵于针叶内时,对其在已着卵针叶附近的针叶上产卵的同种雌蜂会产生驱避作用[14]。而楚雄腮扁叶蜂已交尾雌蜂是否可被正在产卵的雌蜂或者其新卵的挥发物所引诱而聚集产卵,是一个值得探究的问题。为此,我们对楚雄腮扁叶蜂已交尾雌蜂的趋向选择进行了初步的室内测试。
1 材料与方法
1.1 试验材料
1.1.1 供试昆虫
供试昆虫采集于云南省昆明市寻甸回族彝族自治县河口镇北大营。在楚雄腮扁叶蜂即将羽化出土的季节(5月中旬),踏查松林内是否有楚雄腮扁叶蜂危害。选择树冠下散落有前一年度老熟幼虫下树入土前遗留的虫巢的云南松,在树干周边约1 m的半径范围内,挖掘地面深约20 cm,收集经过越冬即将羽化的楚雄腮扁叶蜂蛹,带回室内。在直径25 cm、高15 cm的透明塑料圆盒中先放入湿润的森林土壤,然后放入从林间采回的浅绿色蛹,待其羽化后,分别将雌、雄虫(雌蜂前胸背板及中胸前盾片、盾片和小盾片为锈红色,而雄蜂则为黑色)放在40 cm×40 cm×40 cm的养蜂笼内,喂以10%的蔗糖水,在自然室温20~27℃、相对湿度60%±10%环境下饲养,备用。
以雌、雄虫数目比1∶2放入养蜂笼内任其充分交尾,获得已交尾雌蜂。进行行为测试时,选取未交尾及交尾后1 h的已交尾雌蜂进行测试。
1.1.2 气味源
以云南松新鲜松针、带卵新鲜松针的二氯甲烷淋洗液、粗提液及未交尾雌蜂、已交尾雌蜂产卵器的二氯甲烷淋洗液为气味源。
1.2 试验方法
1.2.1 气味源的准备
松针淋洗液、带卵松针淋洗液的准备:在野外松林中现场制备。采集新鲜松针和带有新卵(卵呈现油亮的黄色光泽)的松针。先将带卵、不带卵的松针轻轻摘下,不要破坏松针上附着的卵,然后以二氯甲烷淋洗。淋洗时要避免淋洗液接触到松针基部的伤口,以免淋洗液遭到松针内次生代谢物的污染。1束松针(3根松针为1束,1束着卵松针附着约85粒虫卵)以10 μL二氯甲烷溶剂淋洗。
松针粗提液、带卵松针粗提液的准备:采用同时蒸馏萃取法(SDE)提取针叶挥发油。将带卵和未带卵的新鲜松针用医用小剪刀剪成长约2 mm的碎屑,各称取200 g放入同时蒸馏萃取装置内,以50 mL二氯甲烷为萃取剂。同时蒸馏萃取5 h后,以布氏漏斗抽滤,旋转蒸发仪除去二氯甲烷,剩余萃取溶液30 mL,用无水硫酸钠干燥。
雌蜂产卵器淋洗液的准备:轻轻挤压雌蜂腹部,待抱卵器伸出后,以二氯甲烷淋洗。每头雌蜂以10 μL二氯甲烷溶剂淋洗。
以上淋洗液和提取液于4℃冰箱内保存,备用。
1.2.2 趋向试验
测试雌蜂趋向行为时,淋洗液直接使用,而带卵松针和未带卵松针粗提液需稀释1 000倍后使用,则10 μL溶液约相当于1束松针的提取液。
利用Y型嗅觉仪,参照张同心等[11]的测试方法(单向选择试验)。测试时,在一个球形管内放入滴有10 μL淋洗液的滤纸条(2.0 cm×0.5 cm)作为气味源,在另一个球形管内放入滴有10 μL相应溶剂(二氯甲烷溶液)的滤纸条作为对照。试验时,将试虫放入Y型嗅觉仪主管入口处,用黑色聚乙烯膜覆盖整个嗅觉仪,以免由于趋光性而影响试验结果。启动真空泵,调节Y型嗅觉仪主管内的气流流量为100 L/h。引入试虫2 min后检查试虫趋向反应:如果试虫进入作为气味源的支管或球形管中,记为正趋向;如果试虫进入作为对照的支管或球形管中,记为负趋向;如果试虫停留在Y型管的主管中,则记为无选择。用正趋向试虫数占被测试的试虫总数的百分比来表示被测试样品的活性(趋向率)。每次测试使用同一批从野外采回的经过饲养的楚雄腮扁叶蜂成虫;每个气味源测试5次,每次测试时将10头雌蜂依次试验;每测试5头雌蜂,调换一次气味源。对经过一轮测试的试虫不再使用。
比较不同气味源的引诱作用(双向选择试验)时,则在Y型嗅觉仪的2个球形管内分别放入不同的气味源。
1.3 数据处理
以Excel 2007进行数据处理并作图,以t测验(t-test)进行差异显著性分析。
2 结果与分析
2.1 提取液、淋洗液对楚雄腮扁叶蜂雌蜂活性的单向选择试验
室内测试表明,楚雄腮扁叶蜂的未交尾雌蜂和已交尾雌蜂对松针粗提液和带卵松针粗提液都具有明显的趋向反应(图1~2)。

图1 未交尾雌蜂和已交尾雌蜂对云南松松针粗提液的趋向反应Fig.1 Approach response of unmated and mated females to crude extracts of fresh needles of P.yunnanensis
从图3可以看出,松针淋洗液既没有引起楚雄腮扁叶蜂未交尾雌蜂的趋向反应,也没有引起已交尾雌蜂的趋向反应。带卵松针淋洗液对未交尾雌蜂没有表现出明显的引诱作用,但对已交尾雌蜂的引诱作用明显(图4)。

图2 未交尾雌蜂和已交尾雌蜂对云南松带卵松针粗提液的趋向反应Fig.2 Approach response of unmated and mated females to crude extracts of fresh needles with fresh eggs of P.yunnanensis

图3 未交尾雌蜂和已交尾雌蜂对云南松松针淋洗液的趋向反应Fig.3 Approach response of unmated and mated females to eluent of fresh needles of P.yunnanensis
未交尾雌蜂产卵器的淋洗液对未交尾雌蜂和已交尾雌蜂没有表现出任何的引诱活性(图5)。已交尾雌蜂产卵器的淋洗液对未交尾雌蜂没有引诱活性,但对已交尾雌蜂具有引诱作用(图6)。
归纳单向选择试验(气味源和二氯甲烷)结果,可以得到以下结论:未交尾雌蜂可被松针和带卵松针的粗提液所引诱,但对松针、带卵松针、未交尾雌蜂产卵器及已交尾雌蜂产卵器淋洗液没有做出趋向反应。已交尾雌蜂对松针淋洗液、未交尾雌蜂产卵器淋洗液没有表现出趋向反应,但可被松针粗提液、带卵松针粗提液和淋洗液、已交尾雌蜂产卵器淋洗液所引诱。
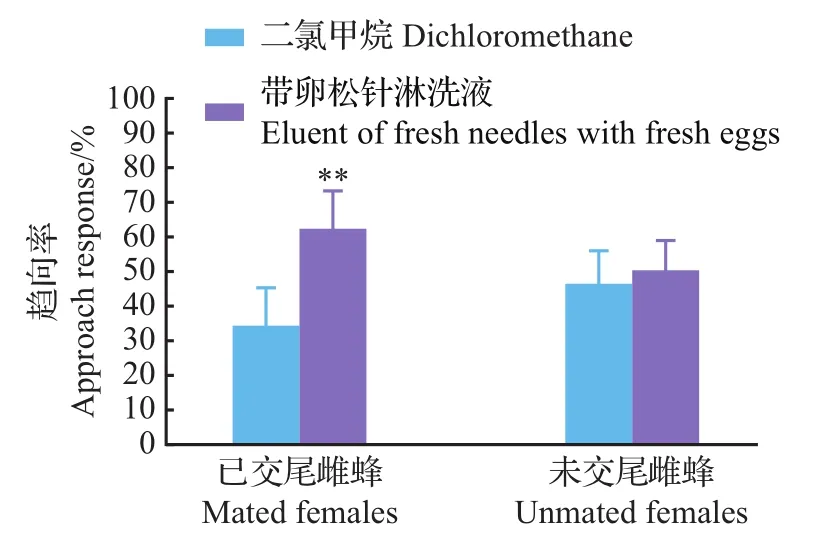
图4 未交尾雌蜂和已交尾雌蜂对云南松带卵松针淋洗液的趋向反应Fig.4 Approach response of unmated and mated females to eluent of fresh needles with fresh eggs of P.yunnanensis

图5 未交尾雌蜂和已交尾雌蜂对未交尾雌蜂产卵器淋洗液的趋向反应Fig.5 Approach response of unmated and mated females to ovipositors’eluent of unmated females

图6 未交尾雌蜂和已交尾雌蜂对已交尾雌蜂产卵器淋洗液的趋向反应Fig.6 Approach response of unmated and mated females to ovipositors’eluent of mated females
2.2 提取液、淋洗液对楚雄腮扁叶蜂雌蜂活性的双向选择试验
单向选择试验表明:未交尾雌蜂、已交尾雌蜂可被松针粗提液、带卵松针粗提液所引诱;此外,已交尾雌蜂还可被带卵松针淋洗液、已交尾雌蜂产卵器的淋洗液所引诱。为了进一步比较楚雄腮扁叶蜂已交尾雌蜂对这些具有引诱活性的气味源的趋向反应是否存在差异,我们进行了双向选择趋向试验。
从图7可以看出,无论是新鲜不带卵松针还是带卵的松针,其粗提液对楚雄腮扁叶蜂已交尾雌蜂的引诱作用远远大于其淋洗液的引诱作用。如图8所示,与松针淋洗液相比较,楚雄腮扁叶蜂已交尾雌蜂选择趋向带卵松针淋洗液和已交尾雌蜂产卵器淋洗液,而对未交尾雌蜂产卵器淋洗液没有做出选择。与未交尾雌蜂产卵器淋洗液相比较,已交尾雌蜂更偏向带卵松针淋洗液;而在带卵松针淋洗液与已交尾雌蜂产卵器淋洗液之间没有做出选择(图9)。产卵器淋洗液之间的比较表明,已交尾雌蜂产卵器淋洗液更有引诱力(图10)。

图7 已交尾雌蜂对新鲜松针、带卵松针的粗提液和淋洗液的趋向反应对比Fig.7 Comparison of approach response of mated females to eluents and crude extracts of fresh needles with or without fresh eggs
3 讨论与结论
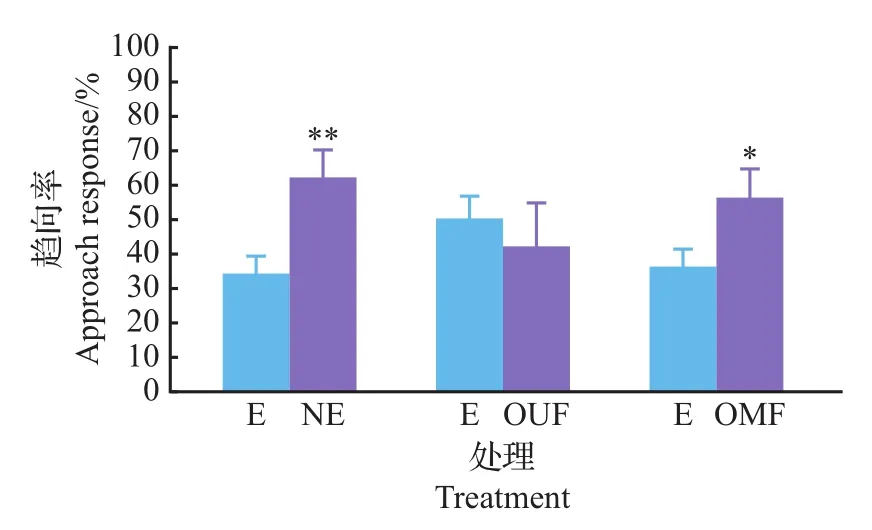
图8 已交尾雌蜂对新鲜松针淋洗液和其他不同淋洗液气味源的趋向反应Fig.8 Approach response of mated females to eluent of fresh needles and other different eluent odors
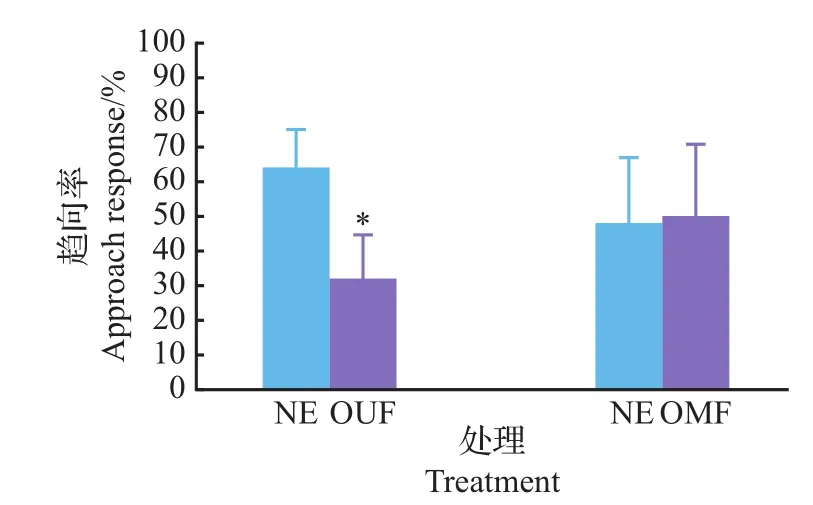
图9 已交尾雌蜂对带卵松针淋洗液与其他淋洗液气味源的趋向反应Fig.9 Approach response of mated females to eluent of fresh needles with fresh eggs and other eluent odors

图10 已交尾雌蜂对未交尾雌蜂和已交尾雌蜂产卵器淋洗液的趋向反应Fig.10 Approach response of mated females to ovipositors’eluent of unmated and mated females
单向选择试验显示,未交尾雌蜂和已交尾雌蜂可被松针粗提液和带卵松针粗提液所引诱,但未对松针淋洗液表现出趋向反应。松针粗提液和淋洗液活性的差别在于其挥发物组成上的差异。植物体内的次生物质是通过表皮渗透到体表或通过腺体结构分泌到体表的,成为该植物的“指纹”信息化合物,进而影响植食性昆虫对产卵寄主的选择性[15-16],说明松针内部的各个单萜烯化合物的相对比例在经过针叶体表蜡质后发生了改变。挥发物的提取方法不同,最后的分析结果差异也很大。如利用水蒸气蒸馏法[17]和利用顶空收集法[18]采集到的云南松针叶挥发物在组成和含量上差异很大;另外,用溶剂提取的粗提液中单萜烯的浓度远远大于破损的针叶在自然状态下释放的单萜烯的浓度。针叶淋洗液中的主要成分是叶面蜡质[19],其单萜烯的浓度极低而可能不被楚雄腮扁叶蜂察觉。由测试结果可以推测,针叶中对楚雄腮扁叶蜂未交尾雌蜂和已交尾雌蜂具有引诱活性的挥发性物质是单萜烯,而不是针叶体表的蜡质。在双向选择试验中,松针、带卵松针粗提液对楚雄腮扁叶蜂已交尾雌蜂的引诱作用远远大于其淋洗液,进一步证实了单萜烯对未交尾、已交尾楚雄腮扁叶蜂雌蜂的引诱作用。然而,已交尾雌蜂可被带卵松针的淋洗液所引诱,由此可以推测,带卵松针淋洗液中引诱已交尾雌蜂的活性物质应该是卵体表附着的产卵分泌物。
未交尾雌蜂对未交尾雌蜂和已交尾雌蜂产卵器淋洗液未表现出明显的趋向反应,而已交尾雌蜂对已交尾雌蜂产卵器淋洗液趋向反应明显。由此可见,在已交尾雌蜂产卵器淋洗液中含有对已交尾雌蜂具有引诱活性的物质。松针淋洗液分别与带卵松针淋洗液、已交尾雌蜂产卵器淋洗液相比较表明,已交尾雌蜂更偏好带卵松针和已交尾雌蜂产卵器淋洗液的气味,试验进一步证实已交尾雌蜂分泌物或者雌蜂新产的卵附着的分泌物中含有引诱性物质。此外,待产卵的雌蜂趋向带卵松针淋洗液而不是未交尾雌蜂产卵器淋洗液,但未在带卵松针淋洗液和已交尾雌蜂产卵器淋洗液之间做出选择,说明卵附着的分泌物和待产卵雌蜂产卵器分泌物具有同等的引诱效应。已交尾雌蜂选择趋向待产卵雌蜂产卵器淋洗液而不是未交尾雌蜂产卵器淋洗液,也说明了两者之间成分的差异。这种差异也说明,楚雄腮扁叶蜂雌蜂交尾后,其生理状态发生了改变,因而其虫体挥发物也发生了变化。
至此,可以得到一幅图景:羽化出土的楚雄腮扁叶蜂雌、雄蜂,首先接触到了林间各种各样的挥发性物质,经过辨别,它们最终选择了来自松树的气味[3];找到其适宜寄主后,雌蜂释放性信息素引诱雄蜂前来交尾[20]。这个交尾后的雌蜂(先驱性雌蜂)选择合适的松树梢并在其针叶上产卵;此时,该雌蜂不再引诱雄蜂,而通过其生殖副腺及其所产新卵挥发出相关的活性物质,召唤更多的待产卵雌蜂聚集到同一个松梢上产卵。这种聚集产卵及此后的幼虫聚集取食,应该是昆虫抵抗寄主植物防御机制的一种生存策略[21]。
综上所述,松针挥发物、生殖副腺分泌物中的挥发性成分(包括新产卵体表的挥发性物质、待产卵雌蜂产卵器分泌物)都对楚雄腮扁叶蜂已交尾雌蜂具有明显的引诱作用。雌蜂的聚集产卵和幼虫的群集危害,是楚雄腮扁叶蜂危害的突出特征。楚雄腮扁叶蜂依赖寄主松树针叶本身释放的挥发物来确定其适宜的产卵寄主,而正在产卵的雌蜂及其新卵的挥发物则在引导同种雌蜂聚集产卵中发挥作用。昆虫产卵时含有调节同种或其他种昆虫产卵行为的化学信息物质,如产卵标记信息素,包括产卵忌避信息素、产卵抑制信息素、产卵刺激信息素等[22]。甘蓝地种蝇(Delia radicum)[23]、沙漠蝗(Schistocerca gregaria)[24-25]卵表面附着的生殖副腺分泌物中含有引诱同种雌成虫聚集产卵的刺激素。膜翅目昆虫的产卵分泌物含有多种化学物质:蚁类产卵分泌物主要是短链烷烃,以及一些萜类、酮类、醇类和酯类;蜜蜂产卵分泌物含有大环内酯、碳氢化合物、类萜、酮类和酯类等;胡蜂产卵分泌物主要包含5~21个碳原子的长链烷烃;寄生蜂产卵分泌物含有丰富的饱和、不饱和烷烃[26]。而楚雄腮扁叶蜂生殖副腺分泌物或新卵体表的具体挥发性引诱物质尚待进一步研究。
有关广腰亚目林木害虫产卵诱导的植物挥发物的研究,大多关注植物应对植食性昆虫产卵引发的直接或间接的防御机制。在为害针叶树的膜翅目食叶害虫中,产卵于针叶内的欧洲赤松叶蜂(D.pini)产卵可诱导寄主欧洲黑松(Pinus nigra)挥发物发生改变[27],欧洲赤松叶蜂在其另一个寄主欧洲赤松(Pinus sylvestris)上产卵,其输卵管分泌物不仅可诱导带卵针叶而且可诱导同一松梢上靠近带卵针叶的其他针叶挥发物发生变化,这种改变了的挥发物对卵寄生的红金姬小蜂(Chrysonotomyia ruforum)具有引诱作用[28-29],其中(E)-β-法尼烯是关键性物质[30]。BEYAERT等认为,松叶蜂产卵引起的寄主松树次生代谢物的变化,是寄主松树自身对即将遭受的幼虫危害而发出的预警性反应[31]。但这种反应是滞后的,松叶蜂产卵72 h后,针叶的光合作用[32]、倍半萜合成酶转录水平[33-34]、挥发物[27,30,35]组成等的变化才达到最大。而楚雄腮扁叶蜂产卵于松树针叶后,诱导改变的针叶挥发物对其聚集产卵产生了什么影响尚不清楚。可以推测,楚雄腮扁叶蜂产卵诱导松树针叶挥发物发生了改变,在这种改变尚未达到驱避或抑制同种雌蜂产卵的效应之前,在一定时间内反而对同种产卵雌蜂具有诱集作用,但这个推测还需要进一步的试验验证。
猜你喜欢
——育王批次和场地的安排
蜜蜂杂志(2023年9期)2023-11-26 23:52:55
环境昆虫学报(2023年2期)2023-05-27 14:00:56
中国蜂业(2021年7期)2021-08-02 01:05:20
山西林业(2021年2期)2021-07-21 07:29:36
昆虫学报(2021年5期)2021-07-07 03:25:32
山东林业科技(2019年2期)2019-06-03 10:11:02
第二课堂(初中版)(2018年7期)2018-09-08 02:54:40
河北大学学报(自然科学版)(2017年2期)2017-04-26 05:26:26
中国蜂业(2016年9期)2016-01-30 06:04:45
生态学报(2013年14期)2013-09-07 02:53:44
