
齐卡兔瘦蛋白基因在不同组织中的表达水平与肉质指标的关联性
2019-09-19 03:12:04罗刚王杰陈婷婷刘林海赖松家
浙江大学学报(农业与生命科学版) 2019年4期
罗刚,王杰,陈婷婷,刘林海,赖松家
(四川农业大学动物遗传育种研究所,成都 611130)
随着生活水平的不断提高,人们的消费价值观也发生了很大的变化,在追求营养和健康的同时,食用动物的肉质和口感也成了人们新的关注点。兔肉是一种营养价值很高的肉食品,其高蛋白、低脂肪的特性使其具有“保健肉”的美称。截至目前,与兔肉品质有关的研究尚未见报道;而在其他动物上的研究表明:pH值是反映猪肉生化变化的重要指标,对猪肉的颜色、嫩度、保水性和肌纤维水分分布[1]有显著影响,pH值越高,蒸煮损失越小[2];对羊肉研究表明,烹调损失越小,出品率越高,肌肉系水力也会越好[3];同时,系水力越高,肉质越好在猪肉中也有报道[4]。滴水损失是衡量系水力高低的指标之一,肌内脂肪(intramuscular fat,IMF)含量与猪肉的嫩度、多汁性和风味高度相关[5]。瘦蛋白基因(leptin)参与调节摄食、基础代谢和胴体脂肪含量,在维持体质量方面起着重要作用。猪血清中瘦蛋白浓度大小与脂肪组织的沉积有关[6];血清瘦蛋白浓度与牛胴体的大理石花纹、背膘厚[7-8],猪的心肾脂肪含量、肌内脂肪含量[9],绵羊的脂肪沉淀和肉品质呈极显著相关性[7-10]。张志宏等的研究表明,瘦蛋白水平跟猪脂肪leptinmRNA表达量呈极显著负相关(P<0.01);背膘厚、眼肌面积和肌内脂肪含量与leptinmRNA表达量极显著相关(P<0.01)[9]。总之,近几年的研究结果表明,leptinmRNA表达量与肉质关系密切,可作为评定肉质的特殊指标,但未见leptin基因在家兔肉质方面的研究报道。本实验研究了家兔的不同组织leptin基因的表达量,分析了其与肌内脂肪、熟肉率、pH值、滴水损失等肉质指标的相关性,以期为家兔的肉质育种提供理论依据。
1 材料与方法
1.1 实验材料
选择四川农业大学雅安校区兔场健康无病的齐卡兔[齐卡大型新西兰白兔(N系)]作为实验动物,分别选取35、70、84和91日龄4个阶段的公、母兔各6只,共计48只。在无菌条件下分别采集左后腿肌组织、左肾周脂肪组织和皮下脂肪组织,立即投入液氮中保存,以备提取总RNA。采集4个日龄阶段腿肌组织各5份,其中:1份保存在-20℃环境下以测定肌内脂肪含量,2份用来测定pH值,其他2份保存在4℃环境下以测定熟肉率和滴水损失。所有实验操作都严格按照四川农业大学实验动物操作规范与福利管理委员会制定的《实验动物操作规范》(川农大校发〔2014〕18号)进行。
1.2 实验方法
1.2.1 pH值的测定
根据BLASCO等[5]的方法,分别在公、母兔宰后45 min和24 h采用便携式可插入pH计测定其pH值。
1.2.2 熟肉率的测定
称取10 g左右(记为mm1)的肌肉在沸水中蒸煮30 min,然后挂到阴凉处,30 min后称量并记为mm2。
熟肉率=mm2/mm1×100%.
1.2.3 滴水损失的测定
测量前将肉样称量(记为md1),然后用金属钩将肉样悬吊,外套一个自封袋,袋口系紧;袋内留有足够空间以接纳肉样渗出的水滴。在4℃冰箱中悬挂24 h后除去塑料袋,用滤纸吸去肉样表面水分并称量(记为md2),计算肉样质量减少的百分率,即滴水损失。
滴水损失=(md1-md2)/md1×100%.
1.2.4 肌内脂肪的测定
采用索氏抽提法,参照GB/T 50096-2003《食品中脂肪的测定》测定肌肉脂肪含量。将肉样从冰箱取出,放于4℃环境下解冻12 h后,剁成肉末,然后放到105℃烘箱中脱水至恒量,冷却,粉碎。精确称取1.0 g经研细干燥的样品(记为mf),无损地移入滤纸筒内,把滤纸和样品放入105℃烘箱中脱水至恒量,称量二者的总质量(记为mf1),然后将滤纸筒放入索氏提取器的提取筒内,由冷凝管的上端加入无水乙醚,至接受器内的无水乙醚量为瓶体积的2/3时,于水浴锅内加热,回流提取6 h,最后,取下接受器,把滤纸筒放到105℃烘箱中烘干至恒量,冷却后称量(记为mf2)。

1.2.5 总RNA的提取及其纯度检测
用重庆拓世众和生物技术有限公司的Magen试剂盒提取总RNA,通过1.5%琼脂糖凝胶电泳检测RNA的完整性,用NanoDrop 2000微量紫外分光光度计检测其浓度,用D260nm/D280nm和D260nm/D230nm的比值鉴定样品纯度。
1.2.6 荧光定量聚合酶链式反应
用生工生物工程(上海)股份有限公司生产的反转录试剂盒(PrimeScrip RT reagent kit)把总RNA合成cDNA第一链,然后用以上cDNA为模板,用荧光定量试剂盒SYBR®Prime ScriptTMⅡ[生工生物工程(上海)股份有限公司]进行荧光定量分析,总反应体系为 10 μL,其中:cDNA 模板 1 μL,SYBR®Prime Ex TapTMⅡ 5 μL,上、下游引物各 0.4 μL,无RNase dH2O 3.2 μL。反应条件:95 ℃预变性30 s;95℃变性10 s,61.4℃退火30 s;熔解曲线温度从65℃上升到95℃,升温速率为0.5℃/s,39个循环。对所有的样本重复测定3次并在每次实验时设立对照组,根据GenBank数据库中家兔基因leptin和βactin的编码区序列(coding sequences,CDs),用Primer Premier 5.0设计荧光定量聚合酶链式反应(quantitative real-time fluorescence polymerase chain reaction,qRT-PCR)引物,并由生工生物工程(上海)股份有限公司合成;引物序列信息见表1。

表1 引物序列信息Table 1 Information of primer sequences
1.3 数据分析
以β-actin基因作为内参基因,采用2-△△CT法测定leptin基因的相对表达量,利用SAS 9.2软件中GLM过程进行单因素方差分析,用邓肯法进行多重比较,结果以平均值±标准差表示,并利用CORR过程计算相关系数和检验相关系数的显著性。
2 结果与分析
2.1 leptin基因在各组织中表达量的变化规律
由图1可知,当齐卡兔为35日龄时,leptin基因在公、母兔的肾周脂肪、皮下脂肪和腿肌中的表达量都没有显著差异。在肾周脂肪中,70、84和91日龄母兔leptin基因的表达量都极显著高于公兔的(P<0.01)(图1A)。在皮下脂肪中,91日龄公兔leptin基因表达量极显著高于母兔的(P<0.01),70和84日龄公兔leptin基因表达量显著高于母兔的(P<0.05)(图1B)。在腿肌中,70和84日龄母兔leptin基因表达量显著高于公兔的(P<0.05)(图1C)。综上表明,在不同日龄的3个组织中,肾周脂肪和腿肌中leptin基因的表达量在母兔中高于公兔,在皮下脂肪中leptin基因的表达量在公兔中高于母兔。
2.2 不同日龄齐卡兔腿肌的肉质变化规律
由表2可知,随着齐卡兔日龄的增大,熟肉率和肌内脂肪含量逐渐增大,而滴水损失逐渐减小。其中:70日龄的齐卡兔肌内脂肪含量极显著高于35日龄的(P<0.01),而滴水损失极显著低于35日龄的(P<0.01);84日龄的齐卡兔肌内脂肪含量极显著高于70日龄的(P<0.01),但滴水损失极显著低于70日龄的(P<0.01);91日龄齐卡兔的滴水损失和84日龄齐卡兔的差异不显著,但91日龄的齐卡兔肌内脂肪含量显著高于84日龄的(P<0.05);70日龄和35日龄公兔的熟肉率差异不显著,但70日龄母兔的熟肉率显著高于35日龄的(P<0.05),84日龄的所有家兔的熟肉率极显著高于70日龄的(P<0.01),91日龄的极显著高于84日龄的(P<0.01)。
由表3可知,屠宰后45 min,齐卡兔的pH值随着日龄的增大而增大;屠宰后24 h,pH值先增大后减小。84日龄是屠宰后45 min和24 h时pH值差值最小的时间段。
总之,肌内脂肪、滴水损失和pH值在91日龄和84日龄的差异都比较小,且91日龄屠宰后45 min和24 h这2个时间点的pH值差值太大会影响家兔肉质,因此,建议家兔屠宰的合理时间为84日龄。
2.3 齐卡兔leptin基因表达量与肉质性状的相关性分析
由表4可知:84日龄母兔皮下脂肪中leptin基因的表达量和腿肌中肌内脂肪含量的相关系数为0.95,呈极显著正相关(P<0.01),与熟肉率的相关系数为0.83,呈显著正相关(P<0.05),与屠宰后24 h的pH值的相关系数为0.67,呈显著正相关(P<0.05)。公兔熟肉率与皮下脂肪、肾周脂肪中leptin基因表达量的相关系数分别为-0.90和-0.89,呈极显著负相关(P<0.01);公兔腿肌在屠宰后24 h的pH值与腿肌中leptin基因的表达量呈极显著负相关(P<0.01),相关系数为-0.85。
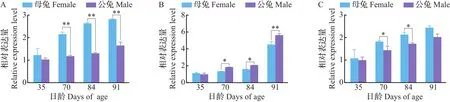
A.肾周脂肪中leptin基因的表达量;B.皮下脂肪中leptin基因的表达量;C.腿肌中leptin基因的表达量。*表示在P<0.05水平差异有统计学意义;**表示在P<0.01水平差异有高度统计学意义。A.Expression level of leptin gene in perirenal fat;B.Expression level of leptin gene in subcutaneous fat;C.Expression level of leptin gene in leg muscle.Single asterisk(*)indicates statistically significant differences at the 0.05 probability level;double asterisks(**)indicate statistically highly significant differences at the 0.01 probability level.
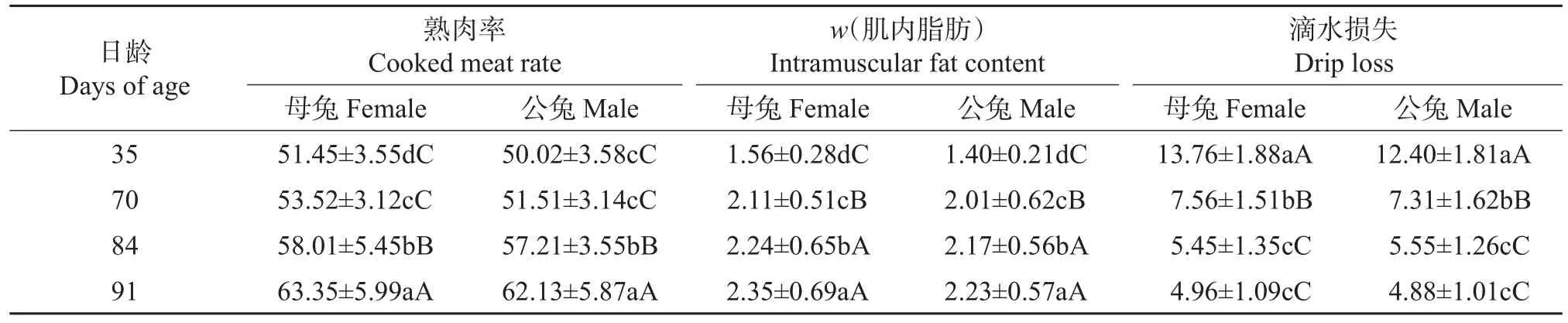
表2 不同日龄齐卡兔的熟肉率、肌内脂肪含量和滴水损失Table 2 Cooked meat rate,intramuscular fat content and drip loss of Zika rabbits at different days of age %

表3 不同日龄齐卡兔屠宰后肌肉的pH值Table 3 pH values of muscle after slaughter of Zika rabbits at different days of age
3 讨论
瘦蛋白作为一种脂肪细胞分泌因子,在调控脂肪沉积与代谢方面发挥着重要作用。研究表明,血清瘦蛋白含量与体内脂肪含量呈正比:当脂肪含量增加时,脂肪组织瘦蛋白分泌会增多,其在组织中的表达量也会增加[11]。在本研究中,随着齐卡兔日龄的增加,leptin基因的表达量在各组织中均升高,说明家兔在生长过程中,肌肉和脂肪组织中的脂肪含量都在提高,与KOLSTAD在猪生长发育过程中的研究结果一致,即在动物的生长发育阶段,其体内总的脂肪含量随着其年龄的增加逐渐增加[12],且leptin基因表达量在肾周脂肪和腿肌中母兔高于公兔,在皮下脂肪中公兔高于母兔。
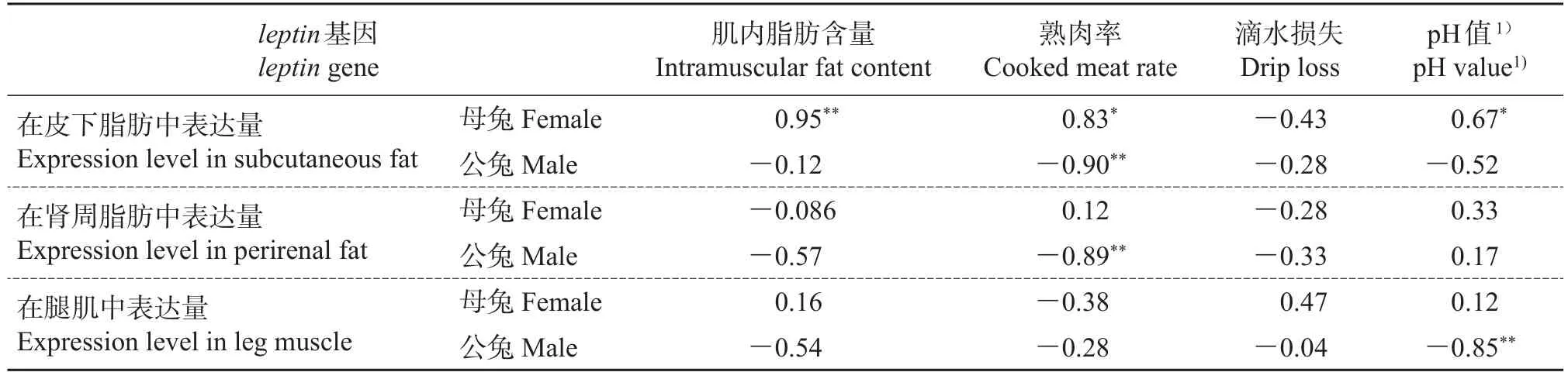
表4 84日龄齐卡兔leptin基因表达水平与肉质性状的相关性分析Table 4 Correlation analysis of expression levels of leptin gene with meat quality traits in the 84 day-old Zika rabbit
瘦蛋白可以通过胰腺、肝、骨骼肌调节糖和脂肪酸代谢,同时还作用于下丘脑的多个神经核团,影响人和动物的摄食量、能量平衡、脂肪沉积[13]。瘦蛋白既可以促进三酰甘油的分解,又可以抑制脂肪酸合成酶的表达,还可以通过增加能量消耗来介导乙酰辅酶A羧化酶基因的表达,从而抑制脂肪的生成[14]。肌内脂肪是重要的风味前体物质。研究表明:脂类物质的氧化是肉香味形成的重要途径,通过加热,脂质发生热降解和氧化降解,形成不饱和羰基化合物(醛和酮),使肉呈特殊香味[15]。在本研究中,齐卡兔leptin基因的表达量在皮下脂肪中前期增长较慢,但在肾周脂肪中前期增长较快,这与KOUBA等在猪的研究上得到的皮下脂肪发育比较缓慢、肾周脂肪生长比较迅速的结果[16]相一致。母兔的肌内脂肪含量和皮下脂肪中leptin基因的表达量呈极显著正相关(r=0.95,P<0.01),说明leptin基因对肌内脂肪有很大的影响。屠宰后pH值迅速下降,呈弱酸性状态,这是肌糖原无氧酵解和乳酸积累的结果。在本研究中,母兔皮下脂肪leptin基因的表达量与屠宰后24 h腿肌的pH值呈显著正相关(r=0.67,P<0.05)。贾小翠等的实验表明,肉鸡宰后pH值的下降会导致肌肉系水能力降低[17];郑云林等研究表明,系水力越低,猪肉质越差[4]。在本研究中,屠宰后24 h的pH值越高肉质越好的结果与上述研究相一致。蒸煮损失与系水力紧密相关,对出肉率有很大影响[18];系水力越高,熟肉率越高,从而可以更好地改善肉的嫩度、风味、滋味、外观和多汁性[19]。
综上所述,齐卡兔的性别和日龄对leptin基因的表达影响显著。母兔的肾周脂肪和腿肌中leptin基因的表达量高于公兔,而公兔皮下脂肪中leptin表达量高于母兔。并且,本研究表明:84日龄家兔pH值在屠宰后下降速度最慢,所以84日龄家兔的肉质最好,是齐卡兔较为适宜的屠宰日龄;此外,母兔熟肉率和皮下脂肪中leptin基因表达量呈显著正相关(r=0.83,P<0.05),且母兔皮下脂肪leptin基因的表达量与屠宰后24 h腿肌的pH值呈显著正相关(r=0.67,P<0.05);公兔熟肉率与皮下脂肪和肾周脂肪中leptin基因的表达量呈极显著负相关(r=-0.90,P<0.01;r=-0.89,P<0.01)。说明leptin基因可能参与家兔肌肉的生长发育过程,可以作为研究家兔肉质的一个候选基因,从而为采用分子遗传标记技术和开展肉兔的遗传改良提供参考。
猜你喜欢
食品安全导刊(2021年21期)2021-08-30 08:21:44
艺术科技(2021年19期)2021-03-13 14:23:13
农村百事通(2020年1期)2020-03-31 03:00:50
重庆医学(2019年22期)2019-12-03 06:33:30
食品安全导刊·下旬刊(2019年4期)2019-10-21 21:58:02
中国医科大学学报(2018年7期)2018-07-06 02:18:04
粮油与饲料科技(2017年3期)2017-03-07 14:24:28
兽医导刊(2016年6期)2016-05-17 03:50:17
畜牧兽医杂志(2015年4期)2015-08-15 00:45:32
医学理论与实践(2015年8期)2015-02-09 20:32:48
