
我国粤东沙菜属(红藻门 杉藻目)的分类鉴定*
2021-03-05 08:55刘金梅姜晶晶马鑫黄冰心杨楠刘美媛丁兰平
热带海洋学报 2021年1期
刘金梅, 姜晶晶, 马鑫, 黄冰心, 杨楠, 刘美媛, 丁兰平
天津师范大学生命科学学院, 天津动植物抗性重点实验室, 天津 300387
沙菜属Hypnea 是广泛分布在热带和亚热带海岸潮间带基质上的大型红藻(Mshigeni et al, 1994; Geraldino et al, 2009; Ding et al, 2013), 其细胞壁含有丰富的K 型卡拉胶成分, 被广泛应用于食品、化妆品和医药等行业(Knutsen et al, 1995; Najam et al, 2010), 某些物种还具有杀菌(Devi et al, 2014)、抗炎(Bitencourt et al, 2008)、抗氧化(Ghaderi et al, 2017)、镇痛(Souza et al, 2019)等作用, 甚至可作为鱼类饲料的添加剂(Mohammad et al, 2018)。
沙菜属由Lamouroux(1813)建立, 模式种为H. musciformis (Wulfen) J. V. Lamouroux, 该属目前已报道了97 个种(Agardh, 1851; Tanaka, 1941; Geraldino et al, 2009, 2010; Nauer et al, 2014, 2015; de Jesus et al, 2015, 2016), 但仅有61 种得到确认(Guiry, 2020)。其主要特征为: 藻体一般直立或缠结, 圆柱状或扁圆柱状, 分枝多, 并被有刺状小枝, 四分孢子囊生于最末小枝或分枝上, 层形分裂, 囊果球形(Mshigeni, 1978; Womersley, 1994; 夏邦美 等, 1999)。
目前, 国际上沙菜属的研究工作主要集中在海产栽培(Ganesan et al, 2006)、生理生化(de Faveri et al, 2015; Ribeiro et al, 2017)、物候学(phenology) (Caires et al, 2013b; de Jesus et al, 2019a)以及卡拉胶的提取工艺(Castelar, 2016)等方面。然而, 在基础的分类学研究方面还存在许多问题有待解决, 如沙菜属的生殖结构特征(Kylin, 1930; Mshigeni, 1976; Vázquez-Delfín et al, 2016)以及准确的物种鉴定标准(Geraldino et al, 2010)。因表型可塑性(Reis et al, 2003)、详细鉴定特征及地理分布信息的缺乏(Masuda et al, 1997; Nauer et al, 2014), 以及不一致的物种划分标准(Yamagishi et al, 2000)等, 导致同物异名现象时有发生(Mshigeni et al, 1994)。我国沙菜属在分类学方面的研究报道非常少, 仅张峻甫等(1963)、Xia 等(1997)和夏邦美等(1999)报道了8 个种, 丁兰平等(2015)总结为15 种, 但后者仅为名录式报道, 缺乏详细的特征描述及地理分布等信息。因此, 需要在更广泛的地域开展专题性研究, 并对物种的划分标准进行重新界定, 才有可能解决沙菜属的分类难题(Masuda et al, 1997)。
此外, 近年来国际上在沙菜属的科级分类系统划分以及物种的分类地位等方面已经有了较大的变动(Saunders et al, 2004; Geraldino et al, 2009; de Jesus et al, 2016)。沙菜科Hypneaceae 藻体一般丛生, 枝呈圆柱状, 被有疏密不等的刺状小枝; 藻体内部具一中轴, 明显或不明显。四分孢子囊生于最末小枝膨大部位的皮层细胞中; 囊果球形, 突出表面。隶属于该科的物种大多数分布在热带和亚热带地区, 然而有学者将其合并到赤叶藻科Cystocloniaceae 中(Saunders et al, 2004; de Jesus et al, 2019a), 由此也引发了一些争议(Geraldino et al, 2009; Islam et al, 2010; Tuastumban et al, 2017)。为解决这些问题, 开展更深入的研究, 本文利用形态学和解剖学方法对采自广东汕头沿海的部分样品进行了分类鉴定, 鉴定结果将为我国对该水域沙菜属的分类和物种多样性的深入了解提供详细资料。
1 材料与方法
1.1 材料
样品于2013—2014 年采集于广东省汕头沿海, 样品处理包括液浸标本(4%福尔马林固定组织)、腊叶标本和干样品(硅胶中干燥保存), 标本或样品保存编号不变。处理后形成的标本存放于天津师范大学藻类实验室。
1.2 实验方法
1.2.1 外形特征观察
首先观察藻体的外观、质地、色泽、主轴和分枝情况, 以及基部和顶端特征等(Tanaka, 1941; Masuda et al, 1997), 然后再对其进行数据测量及拍照(NikonSMZ25、Nikon D7000)。
1.2.2 内部特征观察
利用冷冻切片技术(LEICA CM 1860 UV 生物冷冻切片机)对浸制样品进行切片, 经苯胺蓝溶液染色, 制成临时装片, 放置于正置荧光显微镜(LEICA DM5000 B)下观察, 并进行数据测量及拍照。镜下观察的内部特征主要包括藻体内皮层细胞、外皮层细胞和髓部细胞的大小、层数及排列方式, 以及是否存在髓部细胞镜状加厚(Tanaka, 1941; Masuda et al, 1997)等。
1.2.3 生殖特征观察
生殖特征主要是四分孢子囊的着生位置、外观、排列方式和直径大小, 以及囊果的着生位置、形状和直径大小等(Tanaka, 1941; Masuda et al, 1997)。
1.2.4 物种鉴定
根据上述各特征的观察结果, 结合相关文献和数据进行比对, 并鉴定物种。
2 结果
2.1 密毛沙菜
Hypnea boergesenii Tanaka, 1941: 233-235; 夏邦美 等, 1999: 78; Wai et al, 2009: 11; Islam et al, 2010: 94-94; Kang et al, 2014: 239-241.
特征描述: 藻体紫红色, 软骨质, 丛生, 高6~10cm; 基部具有根状固着器和盘状固着器, 整个藻体密被小枝(图1a、1b、1e); 藻体顶部逐渐尖细, 或众多小枝密被在顶部, 使其不尖锐(图1c、1d); 制成的腊叶标本不完全附着于纸上。
藻体主轴横切面中央具一中轴, 直径约50~70μm(图 1f); 外围为不规则髓部细胞, 较大, 直径约为120~135μm, 呈近圆形(图1g); 内皮层细胞为近圆形; 表皮层细胞为卵圆形, 直径0.8~10μm(图 1h); 髓部细胞壁上具镜状加厚部分(图1i)。
四分孢子囊枝长0.6~1.25mm, 枝端部尖细或分叉, 有的枝上部可再生四分孢子囊枝, 呈念珠状; 四分孢子囊着生于小枝基部、中部、上部膨大处皮层细胞上(图 1j—l), 层形分裂(图 1m), 长径约28~40μm, 短径约17~28.5μm。囊果近球形, 直径约0.5~0.85mm, 单生或集生(图1n、1o); 囊果被由2~3层细胞组成, 厚约100μm(图1p、1r); 果孢子为卵圆形, 径约18μm(图1q)。

图1 密毛沙菜 a. 藻体外观; b. 藻体基部; c、d: 藻体顶端; e. 盘状固着器; f. 藻体横切面; g. 藻体局部横切面; h. 藻体边缘细胞; i. 髓细胞壁镜状加厚; j—l: 四分孢子囊枝; m. 四分孢子囊; n—o: 囊果; p. 囊果切面观; q. 果孢子; r. 囊果被局部切面观 Fig.1 Hypnea boergesenii: (a) habit sketch of the frond; (b) base of the frond; (c~d) upper portion of the branchlets; (e) disk-shaped holdfast; (f) transaction of the frond; (g) transection of part of the frond; (h) cortical cells; (i) lenticular thickening; (j~l) tetrasporangial branchlets; (m) tetrasporangia; (n~o) cystocarp; (p) longitudinal section of the cystocarp; (q) carpospores; (r) longitudinal section of the pericarp
模式标本产地: 中国台湾。标本采集地: 广东汕头。
国内分布: 海南、广东、中国台湾、浙江。国外分布: 菲律宾、越南、日本。
习性: 生长于低潮带的岩石上。
研究编号: 20131205009A(囊果株; n=5)、20131205009B(四分孢子囊株; n=7)。
2.2 裸干沙菜
Hypnea chordacea Kützing, 1847: 776; Tanaka, 1941: 230-232; Xia et al, 1997: 197; Yoshida, 1998: 737-740; 夏邦美 等, 1999: 83; Kang et al, 2015: 199-201.
特征描述: 藻体紫红色, 软骨质, 丛生, 直立, 高约4~8cm(图2a); 近圆柱形, 单条, 叉状分枝少; 主轴下部常裸露, 藻体中上部被有小枝, 枝端不分叉或分叉(图2c); 基部具有根状固着器(图2b); 制成的腊叶标本不完全附着于纸上。

图2 裸干沙菜 a. 藻体外观; b. 基部; c. 藻体顶端; d. 主轴横切面; e. 藻体局部横切面; f. 边缘细胞; g—i: 四分孢子囊枝; j、k: 四分孢子囊局部切面观; l、m: 囊果外观; n. 囊果纵切面; o. 囊果被局部切面观; p. 果孢子 Fig.2 Hypnea chordacea: (a) habit sketch of the frond; (b) base of the frond; (c) upper portion of the branchlets; (d) transection of the frond; (e) transection of part of the frond; (f) cortical cells; (g~i) tetrasporangial branchlets; (j~k) transection of part of tetrasporangia; (l~m) cystocarp; (n) longitudinal section of the cystocarp; (o) longitudinal section of the pericarp; (p) carpospores
藻体主轴横切面上的中轴由一些近圆形小厚壁细胞组成, 径约27~62μm; 外围是由薄壁细胞组成的髓部, 呈不规则卵圆形或长圆形, 径约65~110μm; 髓部细胞壁无镜状加厚部分(图2d—f)。
四分孢子囊枝长 0.5~1.8mm, 枝端一般尖细, 偶有分叉(图2g—i); 四分孢子囊多数为长卵形, 长径约30~55μm, 短径约18~25μm, 层形分裂, 着生于小枝基部或稍上部位的皮层细胞上, 周围细胞常变态延长(图2j、2k)。囊果生于小枝处, 无柄, 近球形(图2l、2m), 径约0.3~1.1mm, 囊果被由4~5 层细胞组成, 厚约80~105μm(图2n、2o)。果孢子呈卵形或近圆形, 径约20μm(图 2p)。
模式标本产地: 印度尼西亚。标本采集地: 广东汕头。
国内分布: 广东、中国台湾、福建、浙江。国外分布: 毛里求斯、印度尼西亚、夏威夷、阿曼、日本。
习性: 生长于中潮带及低潮带的岩石、基质上。
研究标本编号: 20130528005(囊果株; n=5)、20140422024(n=10)、20140516018(n=15)、20140518023 (n=2)、20140518024(n=4)。
2.3 长枝沙菜
Hypnea charoides Lamouroux, 1813: 44; Tanaka, 1941: 243-245; Yoshida, 1998: 738; 夏邦美 等, 1999: 81-83; Yamagishi et al, 2000: 27–35; Geraldino et al, 2009: 606-616.
特征描述: 藻体紫红色, 亚软骨质, 直立, 高约5.5~11cm, 圆柱形; 基部稍有缠结; 有及顶主干, 径约0.5mm; 分枝细长, 互生(图3a—c); 主干及分枝上被有刺状小枝, 无鹿角状分枝(图3d)。制成的腊叶标本附着于标本纸上。
藻体横切面观中央有一明显的中轴, 径约45~60μm; 外围髓部细胞较大, 径约60~150μm; 内皮层细胞较小, 径约18~26μm(图3e); 表皮层细胞呈卵圆形, 径约8~12μm(图3f); 髓部细胞壁上有时可见镜状加厚部分。
四分孢子囊枝长 0.4~2.5mm, 四分孢子囊长30~52μm, 层形分裂, 着生于小枝基部或中部膨大部位皮层细胞上(图3g、3h)。
模式标本产地: 澳大利亚。标本采集地: 广东汕头。
国内分布: 海南、香港、中国台湾、福建、浙江。国外分布: 新西兰、澳大利亚、马达加斯加岛、毛里求斯、印度尼西亚、坦桑尼亚、马来西亚、斯里兰卡、菲律宾、越南、阿曼、韩国、日本。
习性: 生长于中潮带的岩石、基质上。
研究标本编号: 20131012005(n=14)。
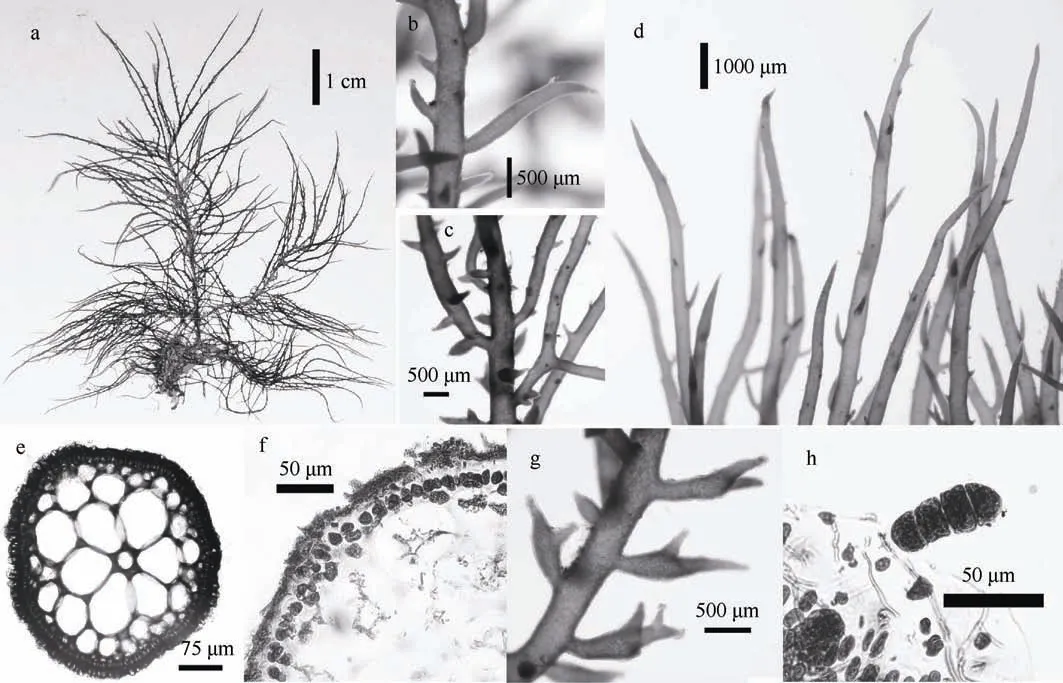
图3 长枝沙菜 a. 藻体外观; b、c: 藻体分枝; d. 藻体顶端; e. 藻体横切面(具镜状加厚部分); f. 藻体边缘细胞; g. 四分孢子囊枝; h. 四分孢子囊 Fig.3 Hypnea charoides: (a) habit sketch of the frond; (b~c) branch of the frond; (d) upper portion of the branchlets; (e) transaction of the frond (with lenticular thickening); (f) cortical cells; (g) tetrasporangial branchlets; (h) tetrasporangia
2.4 冻沙菜
Hypnea japonica Tanaka, 1941: 236-238; 张峻甫 等, 1963: 35-45; 夏邦美 等, 1999: 85-87; Yoshida, 1998: 739, 742; Wai et al, 2009: 12; Tanaka et al, 2011: 593-598.
特征描述: 藻体鲜红色或红褐色, 软骨质, 高5~10cm; 基部具根状固着器, 不规则羽状分枝, 互 生; 及顶主干不明显(图4a、4b、4d); 分枝和小枝顶部形成钩状膨大部分, 缠结在其他藻体上(图4c、4e、4f)。制成的腊叶标本不完全附着于纸上。
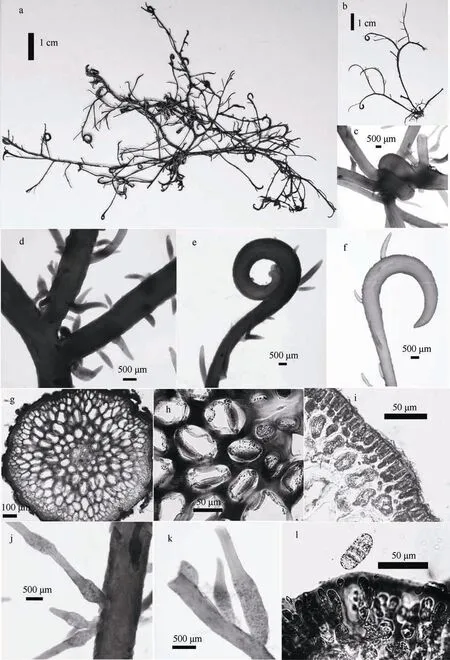
图4 冻沙菜 a、b: 藻体外观; c. 藻体缠结; d. 藻体分枝; e、f: 钩状部分; g. 藻体横切面(具镜状加厚部分); h. 细胞壁镜状加厚部分(箭头所标处); i. 边缘细胞; j、k: 四分孢子囊枝; l. 四分孢子囊 Fig.4 Hypnea japonica: (a~b) habit sketch of the frond; (c) entanglement of the frond; (e~f) hooks of the frond; (g) transaction of the frond; (h) lenticular thickening; (i) cortical cells; (j~k) tetrasporangial branchlets; (l) tetrasporangia
藻体内部横切面的中央具一中轴细胞, 近圆形, 径约25μm, 但此特征在较老的细胞中不明显; 外围由较大的薄壁细胞组成髓部, 呈长卵圆形, 径约 70~80μm(图4g)。内皮层细胞为长圆形或近圆形; 表皮层细胞呈长卵形, 较小, 长 12~18.2μm, 宽4.5~7μm(图4i)。藻体髓层细胞壁大多数具有镜状加厚部分(图4h)。
四分孢子囊枝长1~2mm, 枝端尖细或枝端延长后可再生四分孢子囊枝, 呈念珠状, 易折断(图4j、4k); 四分孢子囊为长卵形, 长径约40~85μm, 短径约20~40μm, 层形分裂(图4l), 着生于小枝基部膨大部位的皮层细胞上。
模式标本产地: 日本。标本采集地: 广东汕头。
国内分布: 海南、广东、香港、福建、中国台湾。国外分布: 印度尼西亚、菲律宾、越南、韩国、日本、俄罗斯。
习性: 可缠结在其他海藻上生长, 或生长于潮下带或低潮带的岩石、珊瑚礁上。
研究标本编号: 20130528001(四分孢子囊植株; n=8)、20140518012(n=4)。
3 讨论
沙菜属的科级分类系统存在争议, 涉及沙菜科Hypneaceae 和赤叶藻科Cystocloniaceae。其中, 赤叶藻科由Kützing 于1843 年建立(Kützing, 1843), 沙菜科由J. Agardh 于1851 年建立(Agardh, 1851)。然而, 这两个科的特征存在交叉现象, 尤其是在沙菜科的物种与赤叶藻科的 Calliblepharis 属之间。 Kylin(1930)最早基于沙菜属和Calliblepharis 属的果孢系结构(procarp structure)将它们归入沙菜科; 之后又基于果孢子囊的差异(沙菜属为单果孢子囊, 而Calliblepharis 属为链状果孢子囊)将它们区分开, 且沙菜属在产孢丝和囊果被之间具有许多吸收丝, 而Calliblepharis 则不存在这样的吸收丝(Kylin, 1932)。尽管它们之间在营养特征上具有许多相同之处, 但Kylin(1932)仍将Calliblepharis 归入赤叶藻科, 仅保留沙菜属在沙菜科中。近年来以分子鉴定为基础的研究认为, 沙菜属为单系类群(Monophyly, 也称单系群, 指包含一个共同祖先和其所有后代的群体)(Geraldino et al, 2010; Nauer et al, 2014, 2015; de Jesus et al, 2015, 2016, 2019b)。也有学者基于SSU(small subunit ribosomal DNA)序列分析了部分杉藻目物种, 认为沙菜科与赤叶藻科应合并为赤叶藻科(Saunders et al, 2004), 但仅选取了沙菜科沙菜属的Hypnea charoides 和 H. ramentacea 两个物种(非沙菜属的模式物种), 其代表性明显不足。此外, 沙菜科和赤叶藻科的卡拉胶组分不同, 前者主要含K 型卡拉胶, 而后者主要含Ι 型卡拉胶(Chiovitti et al 1998), 但这两类卡拉胶在某些杉藻目物种中能共同存在(Knutsen et al 1995)。即使有部分学者认同将沙菜科合并到赤叶藻科, 但仍有许多学者沿用沙菜科作为独立的科, 沙菜属就包含其中(Geraldino et al, 2009; Islam et al, 2010; Miguel et al, 2014; 丁兰平 等, 2015; Tuastumban et al, 2017)。虽然目前分子数据在分类学研究中起到了至关重要的作用, 但生殖结构仍然是区分许多属(杉藻目)的重要鉴定依据(Yang et al, 2015, 2016; Won et al, 2016)。因此, 本文仍然沿用沙菜属为沙菜科的观点。
密毛沙菜H. boergesenii 最早是基于中国台湾的标本命名的(Tanaka, 1941), 之后也被一些学者报道过(Shin et al, 1994; 夏邦美 等, 1999; Islam et al, 2010; Kang et al, 2014)。其藻体一般具有及顶主轴, 整体为密被小枝, 髓部细胞壁上具镜状加厚特征(Tanaka, 1941; 夏邦美 等, 1999; Islam et al, 2010; Kang et al, 2014)。本文研究的样品(编号: 20131205009A、20131205009B)具有明显的主轴, 众多小枝密被于藻体上, 在藻体横切面可明显观察到髓部细胞壁上的镜状加厚, 这些特征与其他文献(Tanaka, 1941; 夏邦美 等, 1999; Islam et al, 2010; Kang et al, 2014)描述的密毛沙菜相一致。据Kang等(2014)的报道, 密毛沙菜的分枝基部具收缢现象, 但这一特征在其他相关文献(Tanaka, 1941; 夏邦美 等, 1999; Islam et al, 2010)中未见报道, 本文的样品也未观察到分枝基部有收缢现象。另外, 编号为20131205009A(囊果株)的藻体枝端由于产生众多密被小枝而不尖细(图1c); 四分孢子囊枝端部具分叉和三分叉, 部分四分孢子囊枝的枝端可再衍生出四分孢子囊枝(图1l), 这些特征在过去的研究中均未见报道。
目前在国际上, 对于密毛沙菜是否为独立种还存在着争议。它与粗糙沙菜H. aspera、鹿角沙菜H. cervicornis 和易曲沙菜H. flexicaulis 等相似, 这几个种在我国也有报道(丁兰平 等, 2015)。粗糙沙菜由Kützing 于1868 年建立(Kützing, 1868), 模式产地为南太平洋的新喀里多尼亚。有学者检查了粗糙沙菜的正模标本(L 941, 97-186/barcode L 0055947)碎片, 发现其藻体上密被小枝, 这与密毛沙菜的特征极其相似, 故认为密毛沙菜是粗糙沙菜的同物异名(Millar et al, 2005), 部分学者也支持这种观点(Payri, 2007; Islam et al, 2010; Nauer et al, 2014)。易曲沙菜是根据日本样品命名的, 其藻体的及顶主轴不明显, 且弯曲, 分枝角度可达150°, 上部分枝呈鹿角状, 髓部细胞壁未见镜状加厚(Yamagishi et al, 2000; Wolf et al, 2011)。密毛沙菜与易曲沙菜的不同之处在于前者没有弯曲的主轴和鹿角状分枝, 分枝角度小, 且髓部细胞壁上具有镜状加厚(Tanaka, 1941; 夏邦美 等, 1999; Islam et al, 2010; Kang et al, 2014), 但基于rbcL 的分子系统发育分析显示两者关系密切(Geraldino et al, 2006)。自2003 年以来, 国际上在涉及粗糙沙菜 H. a s p e r a、密毛沙菜H.boergesenii、鹿角沙菜cervicornis 和易曲沙菜H. flexicaulis 的鉴定方面开展了比较丰富的分子系统发育分析研究, 利用了rbcL(ribulose-1, 5-bisphosphate carboxylase/oxgenase gene)、COI(cytochrome c oxidase subunit I)和UPA(universal plastid amplicon)等3 个分子序列数据(Hommersand et al, 2003; Geraldino et al, 2006, 2009, 2010; Nauer et al, 2014, 2015; de Jesus et al, 2015), 有的学者甚至运用了6 种统计及2 种物种假设方法, 构建了多种类型的系统进化树, 试图探讨这几个物种间的亲缘关系(de Jesus et al, 2016)。综合相关研究的分子分析结果, 这几个物种间或多或少地存在着亲缘关系, 而不同学者和不同方法所得结果不一致, 甚至存在矛盾的情况。因此, 有学者认为精确的物种界定结果还需要采用足够多的样本和开展不同的分子标记分析(de Jesus et al, 2016)。密毛沙菜在形态上与鹿角沙菜和易曲沙菜均具有较大的差异(见表1)。尽管粗糙沙菜与密毛沙菜在形态上差异不大, 但它们的地理分布隔离比较明显, 密毛沙菜主分布于北半球的西北太平洋和印度洋, 而粗糙沙菜则主要分布于南太平洋, 因此也有学者仍视密毛沙菜为独立物种(Kang et al, 2014)。鉴于其分布的生物地理学及形态特征, 本文暂且沿用密毛沙菜为独立种的观点。
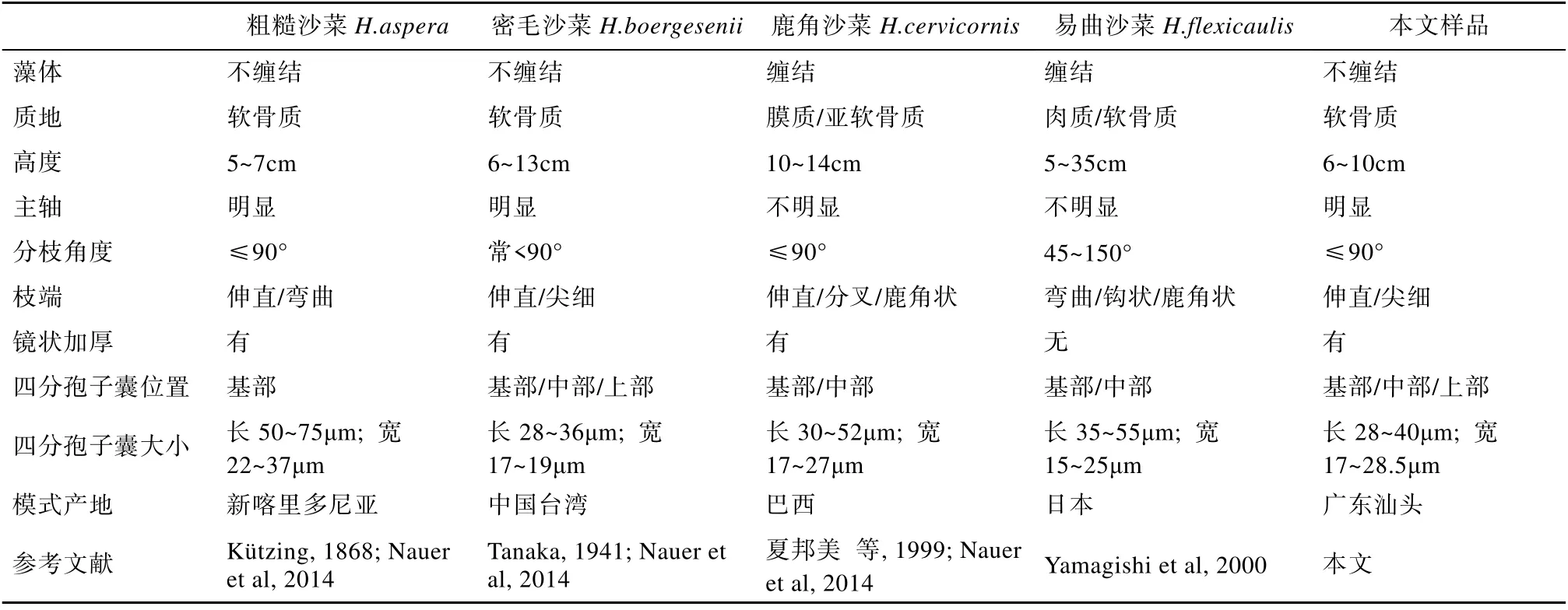
表1 密毛沙菜、鹿角沙菜、易曲沙菜和粗糙沙菜的特征比较 Tab. 1 Comparison of H. boergesenii, H. cervicornis, H. flexicaulis, and H. aspera
裸干沙菜H. chordacea 由Kützing 建立(Kützing, 1847), 主要分布在东亚沿海地区, 如中国、日本等。该种在国内外均较少报道, 其藻体主轴中下部一般裸露, 中轴由许多小厚壁细胞组成, 髓部细胞壁上不具有镜状加厚现象(Xia et al, 1997; Yoshida, 1998; 夏邦美 等, 1999; Kang et al, 2015)。本文样本(编号: 20130528005 、 20140422024 、 20140516018 、20140518023、20140518024)的藻体主轴下部少有小枝, 甚至裸露, 藻体横切面由许多小的厚壁细胞组成中轴, 髓部细胞壁未见镜状加厚情况, 且囊果近球形, 果被4~5 层。其藻体的内、外部特征与大多数学者对裸干沙菜的描述一致(夏邦美 等, 1999)。但Kang 等(2015)所报道的裸干沙菜的外部形态与大多数学者报道的相差较大, 其藻体的分枝程度和小枝的丰富程度均较多, 这种形态上的差异可能是表型可塑性导致的。
长枝沙菜H. charoides 是Lamouroux(1813)最初建立沙菜属的5 个新种之一, 模式产地为澳大利亚。除澳大利亚外, 该种在韩国(Shin et al, 1994)、日本(Yamagishi et al, 2000)、中国(Xia et al, 1997)和马来西亚(Tani et al, 2003)等地也有报道。其藻体具及顶主轴, 分枝互生, 无鹿角状分枝, 小枝向主轴内弯曲, 之后笔直生长(Womersley, 1994; Yamagishi et al, 2000; Geraldino et al, 2006)。本文的样品(编号: 20131012005)藻体具明显的主轴, 分枝互生, 无鹿角状, 小枝细而长, 笔直生长, 这些特征与前人报道中描述的长枝沙菜一致。长枝沙菜和H. valentiae 在形态学上比较相似, 较难区分, 有学者将它们称为“Hypnea charoides-valentiae complex” (Lewmanomont, 1997; Yamagishi et al, 2000)。但根据藻体的分枝角度、主轴的明显程度等特征, 仍可将两者区分开来(Yamagishi et al, 2000 )。长枝沙菜的分枝角度小于H. valentiae, 且小于90°, 具有明显的主轴, 没有鹿角状分枝(Yamagishi et al, 2000)。在一些涉及分子系统发育分析的研究报道中, 它们的遗传距离相对较远(Sargazi et al, 2018; Kang et al, 2018)。有学者认为长枝沙菜(Yamagishi et al, 2000)的生境和形态特征与亚洲沙菜H. asiatica 大致吻合, 应该合并为亚洲沙菜(Geraldino et al, 2009)。此外, 也有学者基于形态特征和cox1 单倍型简约网络分析, 认为长枝沙菜只分布于澳大利亚南部地区, 而不分布在西北太平洋的海藻区系中(Geraldino et al, 2009), 但事实上非洲也曾有报道(Silva et al, 1996)。总的来说, 长枝沙菜是一个分类较为混乱的物种, 其分类地位的确定仍需要更多的证据加以验证。鉴于本文所研究样品(编号: 20131012005)的形态特征及内部结构(图3)与大多数文献中的长枝沙菜描述大体一致, 故暂命名其为长枝沙菜H. charoides。
冻沙菜由Tanaka建立, 模式产地为日本(Tanaka, 1941)。其顶端形成弯曲的钩状结构, 髓部细胞壁具镜状加厚部分(Tanaka, 1941; 夏邦美 等, 1999; Yoshida, 1998)。最初冻沙菜和钩沙菜被认为是近缘物种(Tanaka, 1941), 但后来发现两者的四分孢子囊着生部位不同, 钩沙菜四分孢子囊一般着生在孢囊枝的中部或上部, 而冻沙菜则以着生于囊枝下部为主或稍向上延伸至中部(张峻甫 等, 1963)。本文研究的冻沙菜样品(编号: 20130528001)的四分孢子囊普遍着生于小枝基部, 与其他文献记录相符。据张峻甫等(1963)的报道, 冻沙菜的四分孢子囊见于4—11 月, 也有学者认为四分孢子囊出现的时间为6—7月, 水温为24~26℃(Tanaka et al, 2011)。本文的四分孢子囊植株采集于5 月末, 此时广东汕头沿岸的平均水温约为 25.6 ℃(http://www.tianqihoubao. com/lishi/shantou/month/201305.html), 与其他文献对冻沙菜四分孢子囊的记录大致相符。但本文样品的四分孢子囊枝顶端可再延生出四分孢子囊枝(图4j), 总体长度可达2mm, 这是其他文献中没有提及的现象。总的来说, 本文所研究的冻沙菜样品(编号: 20130528001、20140518012)的内、外部形态特征和四分孢子囊与其他文献中的冻沙菜描述基本一致, 故命名其为冻沙菜H. japonica。
4 沙菜属分种检索表(部分)
检索表是一种为了区分物种而编制的表, 广泛应用于各分类阶元的鉴定。本文通过对沙菜属内的相关物种(密毛沙菜、长枝沙菜、裸干沙菜、冻沙菜)的分类鉴定研究, 认为物种藻体的主轴、小枝形状、枝端形状、四分孢子囊的着生部位和囊果这5 个特征对该属的分类鉴定具有较大价值。但由于沙菜属物种较多, 并且该属是个分类混乱的属, 因此本文只针对研究中的物种编制了相应的检索表, 详细编写如下:
1. 藻体主轴中下部常裸露, 分枝及小枝集中着生于上部.......................................裸干沙菜 H. chordacea
1. 藻体主轴各部位均着生有分枝或小枝................2
2. 枝端弯曲膨大成钩状.....冻沙菜 H. japonica
2. 枝端伸直、不弯曲.......................................3
3. 四分孢子囊着生于小枝基部和中部 ...........................................长枝沙菜 H. charoides
3. 四分孢子囊着生于小枝基部、中部和上部 ........................................密毛沙菜 H. boergesenii
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
广西科学(2022年1期)2022-04-12
东方少年·阅读与作文(2021年5期)2021-06-15
现代农业科技(2020年21期)2020-12-14
上海故事(2020年10期)2020-12-14
水产科学(2019年4期)2019-07-23
生物学通报(2019年7期)2019-07-08
科技创新与应用(2017年20期)2017-07-15
江苏农业科学(2016年4期)2016-06-14
