
适量供锌明显提高平邑甜茶幼苗碳氮吸收和同化效率
2021-03-02 06:32秦嗣军吕德国
植物营养与肥料学报 2021年1期
于 波,秦嗣军,吕德国
(沈阳农业大学园艺学院/辽宁省果树品质发育与调控重点实验室,辽宁沈阳 110866)
氮作为苹果生长发育所必需的重要营养元素,是苹果生长的重要物质基础,对苹果器官建成、物质代谢、产量及品质形成等过程发挥着不可替代的作用[1]。然而,由于果农片面追求高产和大果,我国苹果生产中氮肥过量施用现象普遍,氮肥的过量施用不仅增加了果农的生产成本,降低了氮肥的利用率,还间接造成了土壤酸化、地下水污染以及水体富营养化等一系列生态环境问题[2-3]。因此,提高氮肥利用率,促进氮素高效利用,减少过量施氮带来的资源和环境负担,实现苹果优质丰产是当前苹果生产面临的重要问题[4]。
锌作为植物所必需的微量元素,既可以在植物体内的酶促反应中作为酶的金属组分,又可以作为辅助因子对植物体内酶起到调节、稳定和催化作用,在优化酶结构、提高光合作用、促进糖的运输以及果实产量、品质的调控等方面具有重要作用[5]。相关研究表明,适量供锌可以促进植株根系生长,增强根系活力,提高叶片叶绿素含量,延长叶片功能期,提高净光合速率,促进碳水化合物的运输和分配,提高作物产量[6-8]。过量供锌显著降低了玉米总根长、总根表面积、总根数及总根体积[9],供锌不足则会导致植物体内活性氧 (ROS) 含量增加,细胞代谢紊乱,植物生长发育受阻[10]。氮是植物生长发育的必需营养元素,锌对植株氮素的吸收利用也有显著影响。相关研究表明,增施锌肥能有效促进氮素向“库”的积累与分配,显著提高棉花氮素的积累[11]。锌通过促进根系生长、增大根系表面积、提高根系活力等作用直接促进植株对氮素的吸收[12]。王晓云等[13]研究表明,过量供锌会使得姜苗根系活力、器官全氮量、叶片硝酸还原酶(NR)活性均呈现不同程度的降低,抑制了植株对氮素的吸收与利用。可见,土壤中锌含量的高低会影响植株对氮素的吸收利用。当前,我国苹果生产中锌不足与过量现象并存[14-16]。土壤中锌含量的不适宜可能是导致苹果氮素利用率低的原因之一,迄今为止关于锌对苹果氮素吸收利用及分配影响的研究尚未见报道,相关生理机制有待深入研究。碳氮代谢是植株体内两大主要的代谢过程,两者紧密联系[17]。锌显著影响植株碳代谢[18],而碳代谢与氮代谢在代谢过程与能量水平上互相影响,锌对氮素的吸收利用的影响可能与锌对碳代谢影响有关。因此本研究以苹果生产中常用的砧木平邑甜茶为试材,发挥其植株整齐度高、根系对环境变化较敏感的特点,结合苹果喜硝[19]特性,应用13C与15N同位素示踪技术,探究不同锌水平下苹果砧木15NO3–-N吸收利用及分配特性的差异,以及13C光合同化物在各器官的积累及分配情况,从植株体内碳氮代谢角度出发,为锌直接或间接影响苹果氮素吸收利用提供有力证据,进而为苹果生产中氮素利用率的提高提供理论参考。
1 材料与方法
1.1 试验设计
试验于2019年3—8月在沈阳农业大学果树科研基地进行。以一年生苹果砧木平邑甜茶幼苗为试材,选取生长均匀一致的幼苗移栽到装满石英砂 (消毒后洗净晾干) 的营养钵中 (盆高 21 cm,直径 21 cm,每盆装石英砂4 kg)。石英砂为普通石英砂 (含SiO290%~99%),规格为 120~150 μm。移栽后缓苗1周,期间浇灌去离子水。在Han等[20]营养液配方的基础上去掉其中Zn2+配制基本营养液 (营养液主要成分如下:KNO31 × 10–3、Ca(NO3)2·4H2O 1.6 × 10–3、CaCl20.162 × 10–3、Mg(NO3)2·6H2O 0.5 × 10–3、MgSO40.356 × 10–3、KH2PO41 × 10–3、CuSO4·5H2O 3.91 ×10–3、MnCl2·4H2O 3.9 × 10–3、H3BO31 × 10–2、EDTAFe-Na 4.4 × 10–5、(NH4)6Mo7O243.3 × 10–5mol/L)。缓苗结束后,先用1/2浓度营养液预处理浇灌一周,之后用完全营养液浇灌。
于5月25日开始浇灌含不同浓度Zn2+营养液进行正式处理,每隔3天浇一次营养液,每次浇营养液前用去离子水冲洗3~5次 (洗净之前锌残留的同时防止盐分累积),再浇灌新的营养液,每盆浇营养液 0.5 L。试验设 5个锌 (Zn2+)浓度处理:0 (Zn0)、2.0 (Zn2)、4.0 (Zn4)、8.0 (Zn8)、16.0 (Zn16) μmol/L,其中 Zn 均由 ZnSO4·7H2O (分析纯) 提供。每个处理重复10盆 (各处理试材均分为两组,一组用于后续同位素标记,一组不进行标记,每组各5盆),每盆5 株 (即,Zn0、Zn2、Zn4、Zn8 和 Zn16 处理单株幼苗累积施入锌质量分别为0、0.13、0.26、0.52、1.04 mg)。每次浇灌营养液时每盆加入 Ca (15NO3)20.01 g(共施入0.1 g分10次标记完成) 用于15N标记[标记所用Ca (15NO3)2为上海化工研究院生产,丰度为10.16%]。同时于正式处理25 天后进行13C标记,标记物选用Ba13CO3(13C丰度为98%),用量为0.2 g/plant。将幼苗与标记物、风扇、还原铁粉一同放入密封标记室内 (标记室由透明薄膜做成),于上午9:30开始标记,开动风扇,然后用注射器向装有Ba13CO3的烧杯中注入一定量的 1 mol/L HCl溶液 (为保证Ba13CO3反应充分,HCl溶液应过量),13C标记时间为4 h。同时另选3盆不进行标记,记作空白对照,各处理分别于标记后24、48、96 h对试材进行破坏性采样,用于13C测定。于处理第30天取样测定幼苗其他指标。
1.2 测定指标和方法
1.2.1 植株叶片光合参数的测定 各处理随机选取长势一致植株各5株,用CIRAS-2型光合仪测定叶片的光合能力。选择幼苗新梢顶部第5~6片叶片,记录净光合速率 (Pn)、气孔导度 (gs) 与胞间CO2浓度 (Ci)。用FMS-2便携脉冲调制式荧光仪测定叶绿素荧光参数。测定Fv/Fm时用密闭式适配器配合暗适应夹暗适应30 min后测定,测定非光化学猝灭系数(qN)、光化学猝灭系数(qP)、电子传递速率(ETR)时将光下的叶片加上暗适应夹 (无需暗适应),然后将密闭式适配器扣在暗适应夹上,设置光源为 800 μmol/(m2·s)。
1.2.2 植株生物量、根系形态与根系活力的测定各处理随机选取5株长势一致幼苗,分成根、茎、叶3部分,放入烘箱于105℃杀青30 min,随后80℃烘干至恒重,进行各器官生物量测定。
每处理另选取3株长势一致幼苗,将根部用去离子水清洗后平铺于透明塑料板上,用WinRHIZO(2007版) 根系分析软件进行扫描分析;同时采用氯化三苯基四氮唑 (TTC) 法测定根系活力,根系活力 [μg/ (g·h)]用四氮唑的还原强度来表示[21]。
1.2.3 酶活性的测定 硝酸还原酶 (NR)、二磷酸核酮糖羧化酶(Rubisco)和蔗糖磷酸合成酶 (SPS) 活性参照赵世杰等[22]的方法测定。
1.2.4 植株15N及13C丰度测定 植株取样后,分为根、茎、叶3部分,在105℃下杀青30 min,80℃烘干至恒重后用不锈钢电磨粉碎过0.25 mm筛,样品15N与13C丰度的测定用MAT-251质谱仪,由中国农业科学院农产品加工研究所完成。
1.2.5 植株各器官锌含量测定 先去除幼苗表面附着的锌 (用自来水冲洗,而后在 Na2-EDTA 20 mmol/L溶液浸泡15 min),后用去离子水反复冲洗并用纸擦干,再将植株分解为根、茎、叶3部分,在105℃下杀青30 min,80℃烘干至恒重后参照Zarcinas等[23]的方法,将样品先用HNO3–HClO4消煮提取,后用SP9-400型原子吸收分光光度计 (PYE公司,英国)测定植株各器官锌含量。
1.3 数据处理
1.3.115N的相关计算公式 氮素吸收征调能力(Ndff) = (植物样品中15N 丰度 % -15N 自然丰度 %) /(肥料中15N 丰度 % -15N 自然丰度 %) × 100%;
器官全氮量 = 器官生物量 × 氮含量;
氮肥利用率 = [Ndff × 器官全氮量 (g)]/施肥量 (g)×100%。
1.3.213C 的相关计算公式
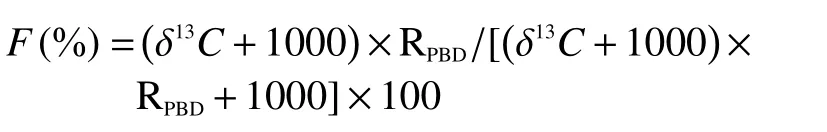
式中:F为13C丰度;RPBD为碳同位素的标准比值(0.0112372)。
各器官13C 积累量:13Ci= (Fi-Fnl) ×TC/100 × 1000式中:13Ci为i器官13C的积累量 (mg);TC为各组分所含的总碳量 (g);Fi与Fnl分别为标记植株与未标记植株13C 丰度 (%)。
用 Microsoft Excel 2010 进行数据处理,用 SPSS 19.0数据处理系统进行差异显著性检验。表中数据为3次重复的平均值 ± 标准差。
2 结果与分析
2.1 不同供锌水平对平邑甜茶生长和根系活力的影响
2.1.1 不同供锌水平下平邑甜茶的生物量 表1显示,不同供锌水平下幼苗各器官生物量均以叶最高、根次之、茎最低。随着供锌水平的提高,平邑甜茶的生物量呈现先升高后降低的趋势,Zn4处理平邑甜茶根、茎、叶生物量最大,其值分别为对照处理的1.43、1.62和1.49倍;随着供锌水平的进一步提高,平邑甜茶各器官生物量呈现下降趋势,Zn16处理平邑甜茶根、茎、叶生物量虽仅为Zn4处理的0.79、0.75和0.78倍,但仍高于Zn0处理。

表1 不同供锌水平下平邑甜茶的生物量 (g/plant)Table 1 Biomass of Malus hupehensis seedlings under different Zn levels
2.1.2 不同供锌水平下平邑甜茶幼苗的根系形态与根系活力 由表2可知,与对照处理相比,供锌处理 (Zn2、Zn4、Zn8、Zn16) 均不同程度地提高了平邑甜茶根系长度、根系总表面积、根尖数,促进了根系的生长。当供锌水平为0~4.0 μmol/L时,随着供锌水平的提高,根系长度、根系总表面积与根尖数逐渐升高,在Zn4处理时达到最高,其值分别为987.31 cm、445.76 cm2和 8742 个,较对照提升了79.83%、68.84%和91.2%。当供锌水平为4.0~16.0 μmol/L时,平邑甜茶根系长度、根系总表面积、根尖数均呈现下降趋势,Zn16处理下平邑甜茶根系长度、根系总表面积以及根尖数仅为Zn4处理的0.67、0.68、0.66倍,但仍高于Zn0处理。可见,当供锌水平为4.0 μmol/L时最有利于平邑甜茶根系的生长。
平邑甜茶幼苗根系活力受供锌水平影响显著(表2)。平邑甜茶幼苗根系活力随着供锌水平的提高呈现先升高后降低的趋势,在Zn4处理时达到最大值 108.6 μg/ (g·h) ,是对照处理的 2.1 倍。可见适宜供锌提高了根系活力,提高了对养分的吸收能力。

表2 不同供锌水平下平邑甜茶幼苗的根系形态与根系活力Table 2 Root architecture parameters and root activity of Malus hupehensis seedlings under different Zn levels
2.2 不同供锌水平下平邑甜茶各器官锌含量
由表3可知,平邑甜茶幼苗各器官锌含量均随着锌水平的升高而上升,幼苗各器官锌含量在Zn16处理下达到最大值。其根系锌含量分别为Zn0、Zn2、Zn4和 Zn8处理的 36.72、15.64、4.34和2.14倍。幼苗各器官内锌含量表现为地下部 > 地上部,当施锌浓度较高时,锌主要在根系中累积,减轻了对地上部的毒害。

表3 不同供锌水平下平邑甜茶各器官锌含量Table 3 Zinc concentration of different organs of Malus hupehensis seedlings under different Zn levels
2.3 不同供锌水平对平邑甜茶碳代谢的影响
2.3.1 不同供锌水平下平邑甜茶的光合参数 由表4可知,平邑甜茶幼苗的光合参数受供锌水平影响显著。Zn0处理下植株净光合速率 (Pn) 与气孔导度 (gs)最低,其值分别为Zn4处理的0.65和0.71倍。当供锌水平为0~4.0 μmol/L时,随着供锌水平的提高,叶片净光合速率 (Pn)、气孔导度 (gs) 逐渐增加,在Zn4处理下达到最大值,其值分别为CK处理的1.55和1.41倍。随着供锌水平的继续提高,叶片净光合速率与气孔导度表现为下降趋势,但仍高于CK处理。不同于叶片净光合速率 (Pn) 与气孔导度 (gs) 变化特征,各处理叶片胞间CO2浓度 (Ci) 表现为随着供锌浓度的提高先下降后升高,Zn4处理叶片胞间CO2浓度 (Ci) 最低。
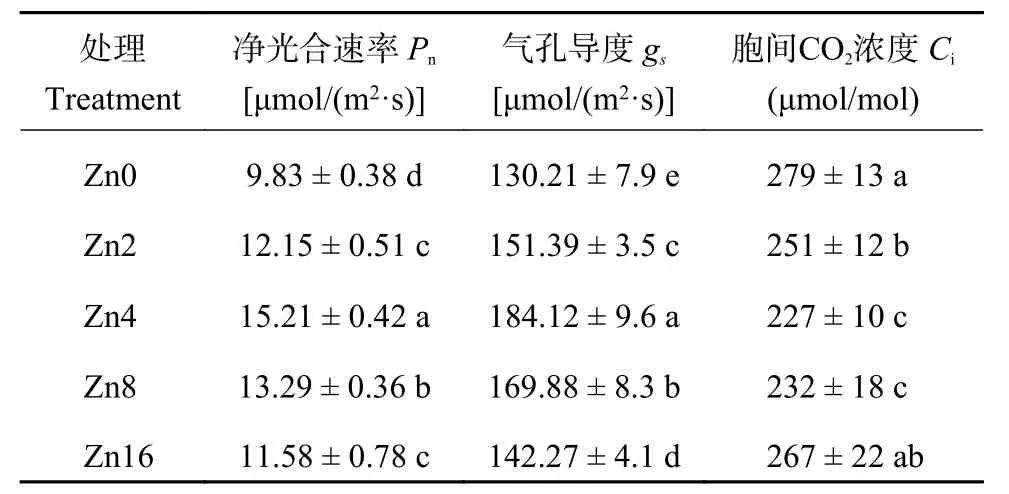
表4 不同供锌水平下平邑甜茶植株的光合参数Table 4 Photosynthesis parameters of Malus hupehensis seedlings under different Zn levels
2.3.2 不同供锌水平下平邑甜茶叶片叶绿素荧光参数
由不同供锌水平对平邑甜茶叶片叶绿素荧光参数的影响(表5)可知,Zn4处理Fv/Fm、ETR以及qP最高,与Zn4处理相比,各处理Fv/Fm、ETR和qP均有不同程度降低,其中以CK处理降幅最大,其值仅为Zn4处理的0.92、0.73和0.88倍。与Fv/Fm、ETR和qP变化趋势相反,Zn0处理qN最大,Zn4处理最低,仅为Zn0处理的0.86倍。可见,不适宜供锌水平均会通过抑制光化学效率和光合电子传递等方式来抑制叶片光合作用。
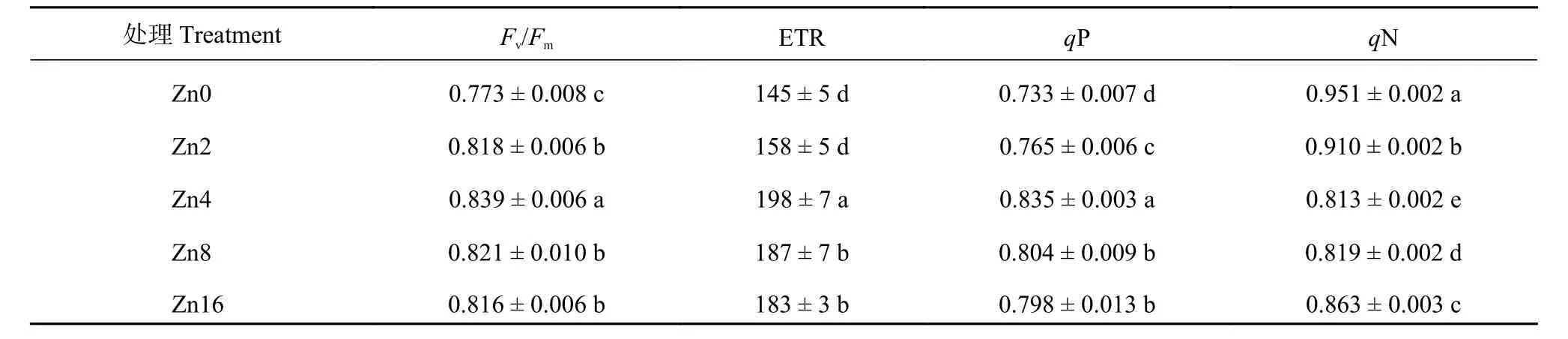
表5 不同供锌水平下平邑甜茶叶片叶绿素荧光参数Table 5 Chlorophyll fluorescence parameters of Malus hupehensis seedlings under different Zn levels
2.3.3 不同供锌水平下平邑甜茶碳代谢酶活性 为了探究供锌水平对碳代谢的影响,测定了叶片中蔗糖磷酸合成酶 (SPS) 以及Rubsico活性。由图1可知,叶片蔗糖磷酸合成酶 (SPS) 和Rubsico活性均在Zn4处理下达到最大值,分别为96.6 mg/(g·h)和8.4 μmol/(g·min),Zn0 处理其值分别为 50.33 mg/(g·h)和 4.28 μmol/(g·min),仅为 Zn4 处理的 0.52 和 0.51倍;而在Zn16处理下则分别下降了28.36%和25.00%。这些结果表明,供锌不足与过量均会显著抑制叶片碳代谢酶活性,且低锌抑制效果较高锌更为明显。
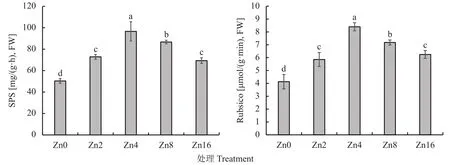
图1 不同供锌水平下平邑甜茶幼苗叶片SPS和Rubsico活性Fig. 1 SPS and Rubsico activity of Malus hupehensis seedlings under different Zn levels
2.3.4 不同供锌水平下平邑甜茶各器官13C积累量
由幼苗各器官13C积累量随时间的变化(表6)可知,13C标记完成后,根与茎中13C积累量随着时间的推移逐渐增加,而叶片中却呈现相反的变化特征。同一取样时间下,幼苗各器官13C积累量随着供锌水平的提高呈现出先升高后降低的趋势,Zn4处理下幼苗各器官13C积累量最高。在13C标记96 h后,Zn4处理幼苗根、茎和叶的13C积累量较对照分别提高了136.8%、63.9%和28.2%,Zn4处理下植株13C 积累量为 1.95 mg,为 Zn0 处理 (1.26 mg)的1.55倍。
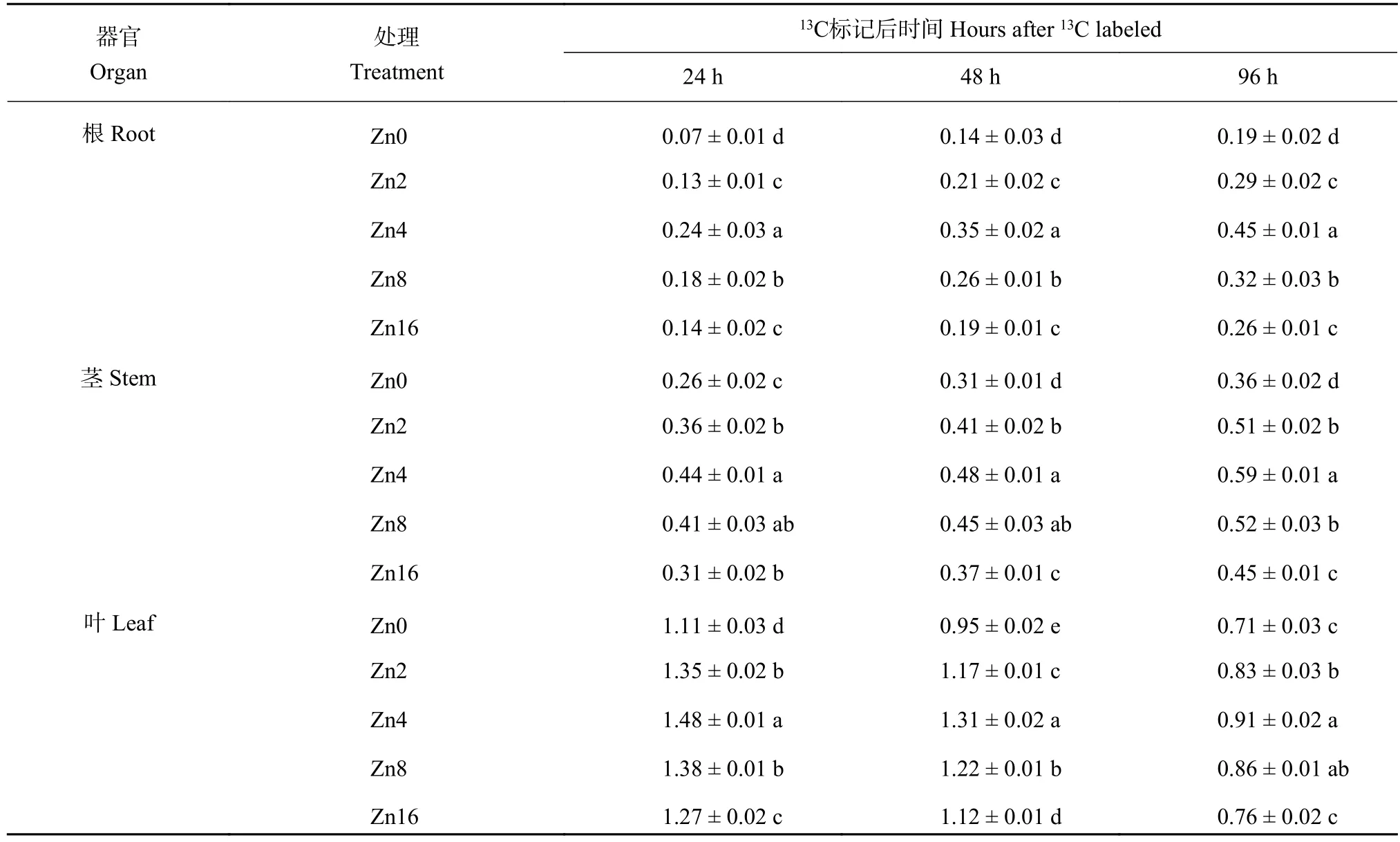
表6 不同供锌水平下平邑甜茶各器官13C积累量 (mg/plant)Table 6 13C accumulation of Malus baccata Borkh. seedlings under different Zn levels
2.3.5 不同供锌水平下平邑甜茶各器官13C分配率
由幼苗各器官13C分配率随时间的变化 (表7)可知,13C标记24 h后,各处理幼苗各器官13C分配率均表现为叶片 > 茎 > 根,随着标记时间的延长,叶片中13C分配率逐渐下降,而根茎中13C分配率则逐渐上升,但仍表现为叶片 > 茎 > 根 (表 7)。13C 标记 96 h后,随着供锌水平的提高,根中13C分配率呈现先升高后降低的趋势,在Zn4处理下达到最大值,为23.1%,为Zn0处理(15.1%)的1.53倍。由此表明,Zn4处理下根系13C竞争能力最强,促进了13C从叶片向根系的运输。

表7 不同供锌水平下平邑甜茶13C分配率 (%)Table 7 13C allocation rate of Malus hupehensis seedlings under different Zn levels
2.4 不同供锌水平对平邑甜茶氮代谢的影响
2.4.1 不同供锌水平下平邑甜茶叶片硝酸还原酶活性
如图2所示,随着供锌水平的增加,幼苗叶片NR活性呈现出先升高后降低的趋势,Zn4处理下达到最大值,为 103.25 μg/ (g·h)。Zn0 处理叶片 NR 活性仅为Zn4处理的0.60倍;在Zn16处理下,叶片NR活性下降了28.96%。
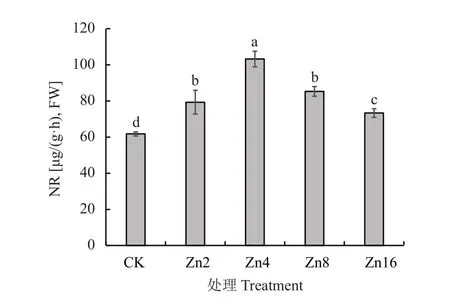
图2 不同供锌水平下平邑甜茶幼苗叶片NR活性Fig. 2 Nitrate reductase activity in leaves of Malus hupehensis seedlings under different Zn levels
2.4.2 不同供锌水平下平邑甜茶15N吸收量、15N利用率以及15N分配率15N同位素示踪结果表明,不同锌水平下苹果幼苗对15N的吸收和利用率存在显著差异。Zn4处理平邑甜茶幼苗的15N吸收量显著高于其他处理,分别是Zn0和Zn16处理的2.19和1.50倍。幼苗15N利用率表现与15N吸收量趋势相同,均以Zn4处理最大、Zn8处理其次、Zn0处理最小。植株全氮量呈现出 Zn4 > Zn8 > Zn16 > Zn2 >Zn0 的趋势 (表 8)。
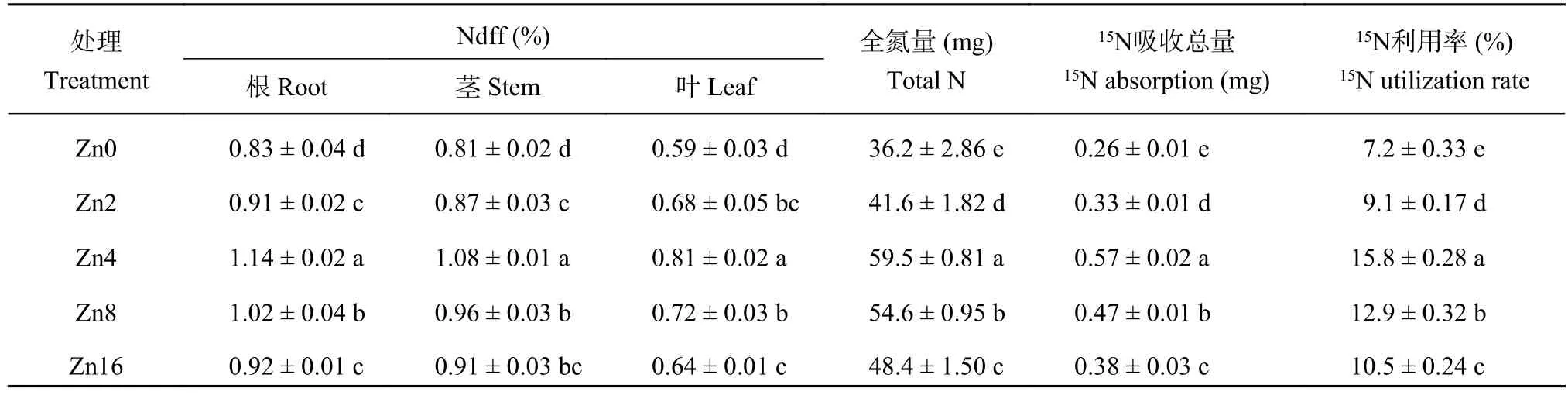
表8 不同供锌水平下平邑甜茶各器官Ndff值和15N吸收量和利用率Table 8 Ndff value,15N absorption and 15N utilization rate of Malus hupehensis seedlings under different Zn levels
图3反映了不同处理下15N在幼苗各器官的分配状况。Zn0和Zn2处理下幼苗吸收的15N主要分布在根部,15N分配率表现为根最大、叶其次、茎最小,而随供锌水平的提高,幼苗吸收的15N向叶片转移增多。Zn4、Zn8和Zn16处理下幼苗的15N分配率均表现为叶 > 根 > 茎。其中,Zn0处理幼苗叶片最低,为36.42%;Zn4处理下幼苗叶片的15N分配率最高,为43.06%,为Zn0处理的1.18倍。由此可见,供锌水平会影响苹果砧木幼苗对氮素的吸收利用和分配,适宜的供锌可促进苹果幼苗对15N的吸收利用,锌不足与锌过量则会抑制15N的吸收利用以及由根系向地上部的转运。

图3 不同供锌水平下平邑甜茶幼苗各器官15N分配率Fig. 3 The 15N distribution of Malus hupehensis seedlings under different Zn levels
3 讨论
3.1 供锌水平对平邑甜茶幼苗根系生长和13C同化及分配的影响
根系是植株生长发育的基础,其生长发育状况可以有效反映植物对养分的吸收能力[24]。锌水平不仅影响了根系的生长,还影响了根系的形态[25]。低锌水平下玉米的根鲜重、根尖数、总根长、总吸收面积和根系活力均明显降低[26]。汪洪等[27]在水稻上研究发现,适宜的锌水平促进了根系生长,提高根系总根长、总表面积,改善了幼苗根系形态,从而有利于水稻生长。王佳[28]研究表明,适当增施锌肥显著促进了冬小麦根系的生长和幼苗地上部的生长,但过量锌肥则对小麦根系有抑制作用。与前人研究结果一致,本研究表明,Zn4处理下幼苗根系生长最好,与Zn4处理相比,Zn0 和Zn16 处理下幼苗根系长度、根系总表面积以及根尖数均显著下降。这可能是因为在缺锌胁迫下根系生长素转运关键基因表达降低[29],生长素含量[30]及其分配发生改变[31],进而影响了根系的生长发育[32];而当供锌水平过高时,植株根系积累大量锌,造成锌中毒现象,使得根系伸长受到抑制,根系生长不良[33]。
光合作用是根系生长发育的基础,根系生长发育及其功能开展所需能量主要来源于叶片的光合作用以及光合同化物[34]。因此叶片光合同化物由叶片向根系的有限运输也解释了为什么不适宜的锌水平影响根系的生长。本研究在前人关于锌对光合作用影响的基础上,采用13C同位素示踪技术,研究锌水平对幼苗叶片光合同化物合成及分配的影响。结果表明,不同锌水平对叶片光合同化物合成及分配影响显著。幼苗13C同化积累量在Zn4处理下达到最高。低锌与高锌处理下幼苗13C同化积累量均显著减少,表明低锌与高锌处理削弱了叶片对碳的固定,抑制了光合产物的合成。13C分配率是各器官对光合同化物竞争能力的体现,根系13C分配率与根系生物量呈显著正相关,根系13C分配率越高,根系生长发育状况越好[35]。进一步分析各处理下幼苗根系13C分配率可知,低锌与高锌处理显著降低了幼苗根系13C分配率。可见,供锌不足或过量在抑制光合产物合成的同时,削弱了根系对13C同化物的竞争力,抑制了光合产物由叶片向根系的运输,降低了根系13C分配率,进而影响了根系的生长。光合作用是叶片光合同化物合成的前提[8]。对幼苗叶片气体交换参数测定发现,低锌与高锌处理下,叶片净光合速率 (Pn) 与气孔导度 (gs) 均显著下降,伴随着净光合速率 (Pn)与气孔导度 (gs) 的下降,胞间 CO2浓度 (Ci) 增高。表明低锌与高锌处理导致的光合速率下降为非气孔限制。进一步分析叶片叶绿素荧光参数可知,低锌和高锌处理下叶片Fv/Fm、qP和ETR均显著低于Zn4(Zn2+4.0 μmol/L) 处理,说明幼苗叶片 PSII反应中心的光化学效率以及电子传递效率在低锌和高锌水平下均受到抑制。Rubisco与SPS是叶片碳代谢过程中的关键酶,直接影响了光合产物的合成及其在植株各器官的分配[36]。Xu等[37]研究认为,叶片Rubisco与SPS活性越高,越有利于同化物向根系的运输。与前人研究结果相似,在本试验中,低锌与高锌处理下叶片Rubisco与SPS活性均显著低于适宜供锌(Zn2+4.0 μmol/L) 处理,这与根系13C 分配率对供锌水平的响应规律一致。可见,低锌和高锌处理削弱了叶片光合作用,抑制碳代谢相关酶活性,阻碍了叶片光合产物的合成及其向根系的运输,影响了根系生长,进而影响了幼苗对氮素的吸收。
砧木为苹果提供了根系,根系生长发育的优劣是苹果氮素高效吸收利用的前提[19,34]。苹果对缺锌极为敏感[15],因此在果园生产中应因地制宜,结合苹果园当地土壤中锌含量水平,相应选择耐性较好的或锌高效型苹果砧木,从而保障苹果对氮素的高效吸收利用。
3.2 供锌水平对平邑甜茶幼苗硝态氮吸收利用及分配的影响
营养元素间的交互作用对改善植物营养元素的吸收、提高肥效等具有重要作用[38]。聂兆君等[39]研究表明,土壤增施锌肥促进了冬小麦对氮素的吸收利用。韩金玲等[40]研究指出,增施锌肥促进了小麦开花前后氮素的吸收积累及向籽粒的运转,提高了小麦籽粒的氮含量。Cakmak等[41]指出,缺锌显著降低了棉花、向日葵和荞麦等植物对硝酸盐的吸收。郭九信等[42]研究发现,锌对小麦茎和籽粒中氮含量及积累量呈现剂量调控效应,即在一定范围内,随着锌水平的提高,茎和籽粒中氮含量及积累量逐渐增加。本研究,采用15N同位素标记技术,分析不同锌水平对平邑甜茶幼苗氮素吸收及分配的影响。15N 示踪结果表明,Zn4 (Zn2+4.0 μmol/L)处理对15N的吸收量与利用率远高于其他处理。这可能是由于 Zn4 (Zn2+4.0 μmol/L) 处理促进了根系生长发育,改善了幼苗的根系形态,提高了根系活力,较好的根系形态与较高的根系活力促进了幼苗对氮的吸收。此外,植株根系吸收氮素以及氮素在植株体内的同化所需的能量与碳骨架均来源于叶片光合作用与碳代谢[43]。因此,Zn2+4.0 μmol/L 处理下植株15N利用率最高可能还与该处理提高了叶片净光合速率以及光合电子传递效率,使得光合碳骨架合成以及能量增加有关。进一步分析15N分配率发现,低锌与高锌处理下叶中15N分配率较低,适宜供锌Zn4 (Zn2+4.0 μmol/L) 处理下叶片中15N 分配率较高,这表明适宜供锌 (Zn2+4.0 μmol/L) 处理在提高根系对氮素吸收的同时,还促进了氮素从地下部向地上部的运输。低锌与高锌处理均抑制了氮素从地下部到地上部的运输,不利于叶片中氮素的积累。这与韩金玲等[40]在小麦上的研究结论相似。Han等[44]研究发现,促进氮素从地下部到地上部的运输,提高叶片中氮素的分配比例,有助于提高叶片对光能的利用效率,增强叶片光合固碳能力,促进植株体内的氮素同化与能量转换,进一步促进了根系对氮素的吸收,提高了氮素利用率。因此,适宜供锌 (Zn2+4.0 μmol/L) 处理下幼苗15N利用率最高,还可能与该处理下叶片中15N分配率较高有关。
氮是植物生长发育过程中需求量最大的元素之一,对植株器官建成、物质合成与代谢等过程发挥着不可替代的作用[19]。有关锌对植株氮代谢的影响在冬小麦[28]、棉花[11]和苦瓜[18]上均有所报道。硝酸还原酶 (NR) 是调节 NO3-同化途径限速的关键酶,在植株体内氮素同化过程中发挥着重要作用,显著影响了植株氮素的吸收与同化[45]。在水稻[46]、苦瓜[18]上的研究均表明,适量增施锌肥有助于提升NR活性,促进了氮素在植株体内的同化利用。本试验结果表明,幼苗各器官锌含量随着供锌水平的提高呈现上升趋势,且均在 Zn16 (Zn2+16.0 μmol/L) 处理下达到最高。与此同时,在一定锌浓度范围内,叶片NR活性随着锌浓度的增加而升高,且在Zn4 (Zn2+4.0 μmol/L) 处理下NR活性达到最高,而此时幼苗各器官锌浓度处于中等水平,随着供锌水平的继续提高,叶片NR活性逐渐下降。可见,适量供锌有助于提高叶片NR活性,而供锌不足与过量均抑制了NR活性,其原因可能是因为锌不足与过量均打破了植株体内锌的平衡状态。结合以上研究结果建议,在果园生产中,果农可在果树需氮关键时期,在氮肥施用的基础上适量配施锌肥来提高氮素利用率,促进氮素高效利用。
综上,本研究分析比较了不同供锌水平下幼苗氮素吸收、利用及分配的差异,有助于探明锌对苹果植株氮素吸收利用的影响机制,为生产上苹果园氮素利用率的提高提供了新思路。为提高结果的可靠性,相关研究还需在大田条件下进一步验证。
4 结论
供锌不足与过量均抑制了幼苗对氮素的吸收利用。适量供锌一方面通过改善叶片光合作用,提高了叶片碳代谢相关酶活性,加强了叶片对碳的固定;提高了根系对光合产物的竞争力,加强了叶片光合同化物向根系的运输,促进根系生长发育的同时,改善了根系形态,提高了根系活力,进而增强了幼苗对氮素的吸收。另一方面,促进了氮素从根系向叶片的运输,同时提高了叶片硝酸还原酶活性,提高了幼苗对氮素的同化利用能力,从而进一步促进了幼苗对氮素的吸收。
猜你喜欢
中国畜牧业(2022年20期)2022-11-19
家庭影院技术(2021年7期)2021-08-14
西北农林科技大学学报(自然科学版)(2021年2期)2021-03-09
江苏农业科学(2019年20期)2019-12-23
中国果业信息(2019年10期)2019-01-05
中学理科园地(2016年6期)2017-01-03
新诗(2015年2期)2015-11-17
现代农业(2015年1期)2015-02-28
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
