
Climate response and radial growth of Pinus tabulaeformis at different altitudes in Qilian Mountains
2021-02-12 03:12LiangJiaoChangLiangQiRuHongXueKeChenXiaoPingLiu
Liang Jiao,ChangLiang Qi,RuHong Xue,Ke Chen,XiaoPing Liu
1.College of Geography and Environmental Science,Northwest Normal University,No.967,Anning East Road,Lanzhou,Gansu 730070,China
2.State Key Laboratory of Earth Surface Processes and Resource Ecology,Beijing Normal University,No.19,Xinjiekouwai Street,Haidian District,Beijing 100875,China
ABSTRACT In order to test whether the relationship between climate and the radial growth of trees is affected by altitude,altitude variability and time stability of climate-influenced radial growth of a dominant conifer,Chinese pine(Pinus tabulaeformis),in the eastern Qilian Mountains were studied against the background of climate change using dendrochronology.Results show that 1)droughts at the end of the growing season of last year and during the early and middle growing season of the current year were the main limiting factors for the radial growth of Chinese pine at two altitude gradients;this was determined by analyzing the relationship between tree-ring width chronologies and climate factors.2)The sensitivity of the radial growth of trees to climate change gradually decreased and was affected more by drought stress at a lower altitude.3)An unstable divergence response was observed in the radial growth at the two altitudes,in response to controlling climatic factors;this observation was based on the moving correlation analysis of growth/climate relationships,and the aggravation of drought stress caused by increasing temperature was the main reason.4)The growth rate of Chinese pine at the two altitudes increased at first and then decreased,as measured by basal area increment (BAI) modeling.Future temperature rises may have significant effects on mountain forest ecosystems in arid and semi-arid regions.Effective management and protection measures should be taken,according to the response patterns of trees to climate change at different altitude gradients.
Keywords:eastern Qilian Mountains;altitude effect;drought stress;divergence response;Pinus tabulaeformis
1 Introduction
Global warming has become an indisputable fact.On a spatial scale,warming trends are not consistent across different regions,with the highest temperature growth rate occurring in the mid-high latitudes of the Northern Hemisphere (IPCC,2013).Great differences in climate change patterns have also been observed,showing a climate characteristic of a transition from warm-dry to warm-wet in northwest China,part of the hinterland of Eurasia (Shiet al.,2007).Moreover,climate change is strongly affecting terrestrial vegetation ecosystems and causing a series of ecological problems,affecting species distribution,forest productivity,biodiversity,plant biological temperature and the global carbon cycle (Lenoiret al.,2008;Kinget al.,2013;Hartl Meieret al.,2014).Forests are considered to have limited adaptability to climate change due to their long life and sensitivity to climate factors.For example,increased drought stress has led to a loss of forest productivity in the eastern Alps and northern Kazakhstan in recent decades (Seidlet al.,2011;Gessneret al.,2013).However,higher temperatures have also accelerated the rapid growth of boreal forests,which is restricted by low temperature conditions (Fritts,1974;Yuet al.,2005).Therefore,forest development is still highly uncertain in the context of climate change,and its impact will depend on the characteristics of different regions and the specific adaptability of species(Elkinet al.,2013).
With the warming of the climate in the second half of the 20th century,sensitivity of the radial growth of trees to climate factors decreased in the middle and high latitudes of the Northern Hemisphere,and this phenomenon is known as the 'divergence problem' (D'Arrigoet al.,1992;Wilmkinget al.,2004).However,numerous studies have found that results relating to response stability of the radial growth of trees to climate change are very complex,which has led to the deepening of studies in this area(Zhanget al.,2009;Jiaoet al.,2015).For example,tree radial growth in the high latitudes of the Northern Hemisphere,specifically in Alaska and the eastern and northern Iberian Peninsula,gradually become less sensitive to temperature with climate warming (Jacobyet al.,1995;Andreuet al.,2007;D 'Arrigoet al.,2008).However,the radial growth ofPicea crassiformisandSibirica ledebin the Qilian and Tianshan Mountains show enhanced responses to temperature with the intensification of drought stress in recent decades (Zhanget al.,2009).Meanwhile,there were no significant differences in the response sensitivity ofLarix gmeliniiKuzen in the European Alps andPicea asperataMast.in northwest China to dominant climate factors (Büntgenet al.,2008;Zhanget al.,2009).Therefore,response stability of the radial growth of trees to climate change is still very uncertain,due to the differing influences of habitat,tree species and age.It is necessary to carry out dynamic research on the tree growth/climate relationships in more regions and among different tree species.
Accurate tree ring dating with high resolution and strong continuity can accurately reflect annual changes and trends in tree growth,providing an important method for studying the impact of climate change on tree growth (Hasenaueret al,1999;Stoffel and Corona,2014).Mountain forest ecosystems are widely regarded as climate change sensors,and the sensitivity of the vertical mountain zone to climate change is much higher than that of the horizontal zone (Malansonet al.,2011;Wanget al.,2015).Therefore,altitude is an important factor affecting the spatio-temporal response results of radial growth to climate change in mountain environments (Wanget al.,2005;Zhanget al.,2015).A large number of studies have shown that the radial growth of trees has an altitude-dependent response to climate factors;that is,the radial growth of trees in high-altitude areas is mainly limited by low temperature,whereas that in low-altitude areas is mainly restricted by precipitation and drought(Frittset al.,1965;Hugheset al.,2003;Tardifet al.,2003;Yuet al.,2007).However,this rule is not applicable to all regions,due to spatial differences of climate in each region (Wanget al.,2005;Lianget al.,2010;Shiet al.,2018).On the one hand,radial growth of trees at high altitude in northwestern Argentina and in arid and semi-arid regions of Central Asia was mainly influenced by precipitation,rather than lower temperature (Moraleset al.,2004;Liuet al.,2006).On the other hand,the radial growth ofPinus taiwanensisat high altitude in Zhejiang Jiulong Mountain and Anhui Guniujiang nature reserve of China was influenced by both temperature and precipitation (Liet al.,2014).In addition,the radial growth of trees in Xinjiang Selimu Basin,in the central Qilian Mountains,shows no difference in altitude response to climate factors (Zenget al.,2012;Qinet al.,2017).Therefore,the study of the response mechanism of tree growth to climate change at differing altitude gradients is helpful to evaluate and predict the developing trends of forest ecosystem in dealing with future climate conditions.
The Qilian Mountains,located in an arid area of northwest China,with their obvious vertical zones of mountain vegetation,are ideal areas to study the response stability of tree growth to climate change along the altitude gradient.Chinese pine (Pinus tabulaeformis) is one of the main tree species in the forests of Qilian Mountain,and it is also a typical conifer species in arid and semi-arid areas,with drought and cold resistance.At present,the dendroclimatology and dendroecology in the Qilian Mountains has mainly focused on the historical climate reconstruction and linear growing-climate relationships (Zhanget al.,2009;Yanget al.,2011;Zhanget al.,2011).However,studies on the nonlinear and divergence response of radial growth to climate factors based at different altitudes have not been perfect or systematic in the eastern Qilian Mountains.Therefore,we took the Chinese pine in the eastern Qilian Mountains as our research object,building two tree-ring width chronologies at high and low altitude gradients,analyzed the limiting factors of the radial growth of Chinese pine at these two different altitudes and discussed the dynamic characteristics of the radial growth response to climate change.Thus,illustrating the ecological adaptability of a dominant coniferous tree in the Qilian Mountains against the background of climate change,we provide a theoretical basis for the protection and sustainable management of forest ecosystems in vulnerable regions.The main purposes of this study are as follows:1) to compare the differences of the main climate factors controlling the radial growth of Chinese pine trees at different altitude gradients;2) to evaluate the response stability of radial growth of trees at different altitudes to climate change;3) to analyze the difference in growth patterns and trends of Chinese pine at different altitudes against the background of climate change.
2 Materials and methods
2.1 Study area
The study area is the Tulu Trench Forest Park(36°40′ N-36°44′ N,102°36′ E-102°45′ E,elevation gradient 2,000-3,500 m),located in the national nature reserve of the Qilian Mountains,with a temperate continental climate (Figure 1).The soil and vegetation resources are abundant,and the region exhibits obvious vertical zonation characteristics.The soil types are subalpine meadow soil,grey brown soil and chestnut soil.The dominant forest species arePinus tabulaeformis(Chinese pine),Sabina przewalsii(Qilian juniper),Picea crassiformis(Qinghai spruce),Picea wilsonii,Populus davidiana,Betula platyphylla(Red birch).

Figure 1 Locations of the sampling site and the nearest meteorological station
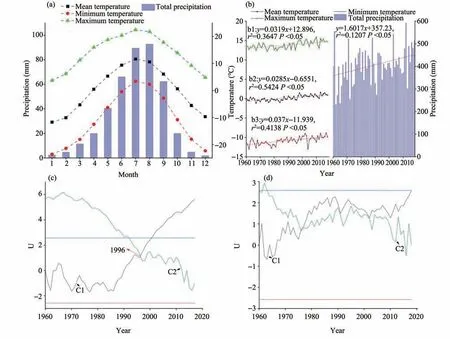
Figure 2 Monthly(a)and interannual(b)variability and Mann-Kendall test(c)annual mean temperature,(d)annual total precipitation;(C1 represents U values for the normal time series,C2 represents U values for the retrograde time series,and the solid lines indicate the value of significance at the 0.01 level)of total precipitation and minimum,mean,and maximum temperatures during 1960-2018
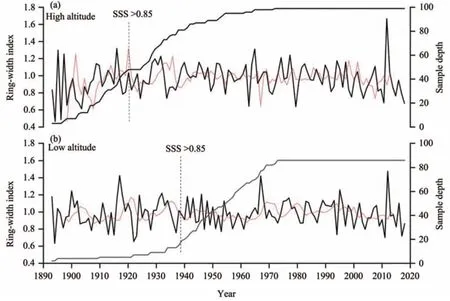
Figure 3 Tree-ring width residual chronologies and sample depth(number of cores)at different altitudes(red line represents 10-year moving average;SSS>0.85 means starting year of EPS reaching or exceeding 0.85)
2.2 Meteorological data
Climate data were taken from the Wushaoling meteorological station (102°87'E,37°20'N,3,045.1 m above sea level,about 50 km from the sampling points).The meteorological data used includes monthly mean,maximum and minimum temperatures and total precipitation during 1960-2018.The radial growth of trees was not only affected by the current year's climatic conditions,but was also restricted by climatic factors of the previous year,so the climate factors from September of last year to October of the current year were selected.
The annual mean temperature is 0.20 ℃,mean minimum temperature is -10.83 ℃and mean maximum temperature is 13.85 ℃,with the frost-free period lasting 125-135 days in the study regions.The mean total precipitation is 419 mm,concentrated from June to September,accounting for more than half of the annual precipitation,and the mean evaporation is 1,542 mm (Zhanget al.,2013).The year-grade change of mean temperature is 0.29 ℃/10a,year-grade change of mean minimum temperature is 0.37 ℃/10a,year-grade change of average maximum temperature is 0.32 ℃/10a and mean change of precipitation is 16.02 mm/10a.There is an obvious raising trend of precipitation and temperature,indicating a change from warm and dry conditions to warm and wet conditions.The annual mean temperature exhibited a sudden increase in 1996,but mean total precipitation did not show any clear sudden variation,according to analysis using the Mann-Kendall method.
2.3 Field sampling and establishment of chronology
Forest community of Chinese pine in the study area is located at an altitude of 2,000-2,300 m.Our sampling sites are located at an altitude of 2,055 m and 2,260 m.The slope is about 20°and the canopy density is small.Individuals with natural growth and less influence by human activities were selected in order to reduce the influence of non-climatic factors on the radial growth of the trees.Two cores were collected from each tree with a 5.15-mm-diameter increment borer.One core was bored parallel to the contour line and the other core was bored orthogonally to the first at breast height (1.3 m above the ground).A total of 100 cores from 50 trees were collected in August 2019.
In the laboratory,tree-ring chronologies were developed using standard dendrochronology techniques(Stokes and Smiley,1968).All cores were air-dried,glued into slotted wood boards,and polished to allow for the visualization of annual rings with 120-,400-,and 600-grit sandpapers.Ring widths were measured using the LINTAB Measurement System with a resolution of 0.001 mm (TM5,Rinntech,Heidelberg,Germany).The qualities of ring width for each species were cross-dated and statistically determined using the COFECHA program (Holmes,1983).To remove age-related and localized disturbance-related trends,raw tree-ring width series were detrended by a negative exponential curve or linear regression.Then,the standard chronologies (STD) and residual chronology(RES) were developed by averaging all individual tree core chronologies with a bi-weight robust mean utilizing the ARSTAN program(Cook,1985).In addition,the residual chronology removed the biological persistence problem in the resulting series and avoided possible "end-effect" bias using autoregressive modeling,which is more suitable for assessing the stability of radial growth responses to climate change(Schneideret al.,2014).Therefore,we finally chose the RES for our analysis of the growth/climate dynamic relationships.
2.4 Dendrochronological parameters analyses
Several dendrochronological parameters were calculated in order to assess the reliability of the chronologies (common period:1960-2018),including MS(mean sensitivity),SD (standard deviation),AC1(first-order serial autocorrelation),R (mean correlation for all series),R1 (mean correlation within trees),R2 (mean correlation between trees),PC1 (variance in the first principal comment),SNR (signal-to-noise ratio),and EPS (expressed population signal) (Table 2).MS characterizes the variability in the ring width index between consecutive years;SD estimates the interannual variation of each series;AC1 detects the effect of the previous year's growth on the current year's growth;R reflects the common signal strength among tree-ring chronologies;PC1 indicates the percentage of variance,explained by the first component in a principal component analysis;SSS is the sample size where the interpreted signal of the sample to the population reaches 0.85.SNR and EPS express the strength of common climatic information among treerings.The chronology was accepted for dendroclimatological analysis if the EPS>0.85(Wigleyet al.,1984).
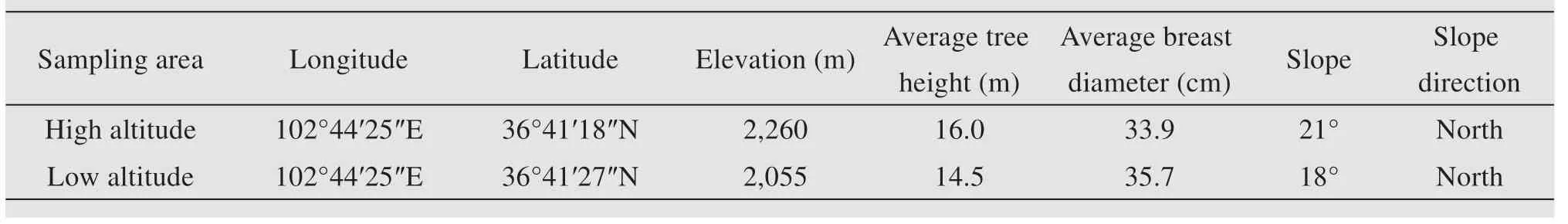
Table 1 Sampling area information

Table 2 Statistical characteristics of chronologies at different altitudes(common period 1960-2018)
2.5 Data processing
In order to determine the main climatic factors affecting the radial growth of trees,the Pearson correlation coefficients between the chronologies and climate factors were calculated using SPSS 25 software.Meanwhile,in order to accurately assess the response stability of radial growth to climate change,the moving correlation between chronologies and climate factors was calculated using DendroClim 2002 software with a fixed year backward sliding window method(moving window:30 years) and bootstrap resampling 1,000 times(Jiaoet al.,2019).
We analyzed the radial growth trend of trees by evaluating the RES chronology and calculating the basal area increment (BAI was calculated from the original chronology).BAI is a better parameter to represent tree growth patterns,because it preserves the original data and eliminates any data conversion biases.The calculation formula of BAI is as follows:
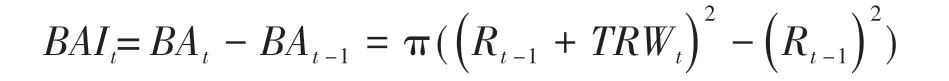
where theBAvariable represents consecutive cross-sectional basal areas,Ris the core length measured for dated tree-rings formed in yeart-1 andTRWis the measured raw width of the tree-ring in yeart.
3 Results
3.1 Comparison of chronology characteristics at different altitudes
Table 2 shows the chronology parameters at high and low altitudes.Both EPS values were greater than 0.85 (high altitude:0.931 and low altitude:0.924) and SNR (high altitude:13.455 and low altitude:5.850)was relatively high,which illustrates that the two chronologies contained strong climate information and were of high quality.At the same time,the SD and MS at high altitudes(0.183 and 0.178,respectively) were higher than those at low altitudes (0.164 and 0.147),indicating that the radial growth at high altitudes was more sensitive to climate factors than that at low altitudes.AC1(0.355)at low altitude was higher than that at high altitude (0.209),indicating that the radial growth at low altitude was more susceptible to the influence of climatic factors in the previous year.
3.2 Analysis of growth/climate relationships at different altitudes
The correlations between chronologies and climatic factors such as minimum,mean and maximum temperatures and total precipitation were analyzed (Figure 4).The results show that the radial growth of trees at the two altitudes was mainly limited by the mean and maximum temperatures in the early and middle growing season,and the radial growth at low altitude was also affected by total precipitation in the middle growing season.
The chronology at high altitude significantly negatively correlated with the mean temperature in September of the previous year (r=-0.300,p<0.05),January (r=-0.296,p<0.05) and July (r=-0.276,p<0.05)of the current year(Figure 4a).The chronology at low altitude had significant negative correlations with the mean temperature of September of the previous year(r=-0.375,p<0.05)and January(r=-0.305,p<0.05),April (r=-0.328,p<0.05) and July (r=-0.353,p<0.01)of the current year(Figure 4a).
The chronology at high altitude significantly negatively correlated with the mean maximum temperature in March (r=-0.312,p<0.05) and July (r=-0.383,p<0.01) of the current year (Figure 4c).Chronology at low altitude significantly negatively correlated with the mean maximum temperature in January (r=-0.288,p<0.05),May (r=-0.302,p<0.05) and July(r=-0.399,p<0.01) of the current year(Figure 4c).
The chronology at low altitude had a significant positive correlation with the total precipitation in July(r=0.267,p>0.05)of the current year(Figure 4d).
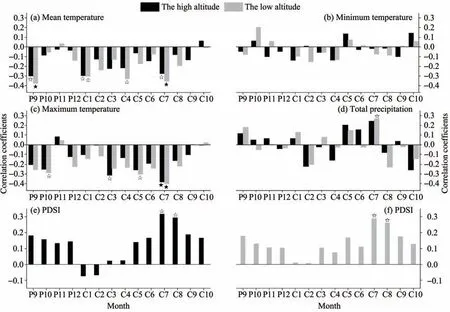
Figure 4 Correlation results of chronologies at different altitudes and climatic factors(black solid five-pointed star reaches 0.01 significance level,black hollow five-pointed star reaches 0.05 significance level;P:previous year,C:current year)
The chronologies at high and low altitudes both significantly positively correlated with the growth season drought index (PDSI),showing the chronology at high altitude significantly positively correlated with July (r=0.317,p<0.05) and August (r=0.294,p <0.05),and that at low altitude significantly and positively correlated with July (r=0.289,p<0.05)and August(r=0.261,p<0.05)(Figures 4e and 4f).
3.3 Evaluation of response stability of radial growth to climate change at different altitudes
The dynamic responses of radial growth to climate change at different altitudes were evaluated by moving correlation analysis (Figure 5).At high altitude,a negative correlation was observed between the chronology and temperatures (mean and maximum)in September of the previous year.Meanwhile,negative correlations between the chronology and temperatures (mean and maximum) in July of the current year were decreasing gradually.At low altitude,positive correlations between the chronology and total precipitation in July of the current year and negative correlations between the chronology and temperatures (mean and maximum) in September of the previous year and July of the current year were increasing gradually.In addition,negative correlations between the chronology and temperatures (mean and maximum) in April of the current year were decreasing gradually.
The results of response stability of radial growth to climate change at different altitudes show the temperature at the end of the growing season of the previous year increased the limiting effect on radial growth of trees at both altitudes,and the drought in the middle of the growing season increased the limiting effect on radial growth of trees at low altitudes.
3.4 Depiction of radial growth trend of trees at different altitudes
The radial growth trend of trees was analyzed by calculating BAI along altitudes (Figure 6).The growth trend at the two altitudes was relatively consistent,with an increase at first,and then a decrease.However,the difference in radial growth patterns between the two altitude gradients was obvious.The radial growth rate at high altitude show an increase before 1968 (1.795 cm2/10a),and then a sharp decline(-1.191 cm2/10a).A similar divergence pattern at lower altitude occurred in 1993,with an upward trend before 1993 (1.563 cm2/10a) and a downward trends after 1993 (-1.356 cm2/10a).Therefore,trees at high altitudes were more sensitive to climate change,showing an earlier downward trend than that of trees at low altitudes.However,the growth at low altitude was under greater drought stress,showing a bigger downward trend than that of trees at high altitudes.
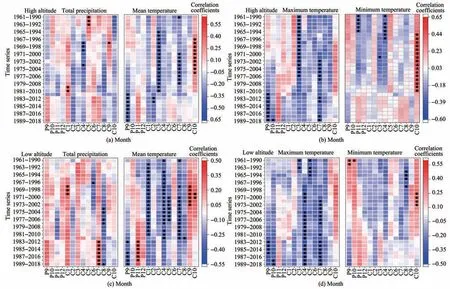
Figure 5 Moving correlation between chronologies and monthly climate factors(total precipitation and minimum,mean and maximum temperatures)at different altitudes(Moving window:30 years,the black solid dots represent significance at the 0.05 level and black solid triangles at 0.01 level;P:previous year,C:current year)
4 Discussions
4.1 Differences in main climatic factors controlling the radial growth of Chinese pine at different altitudes
There has been general warming since the second half of the 20th century in arid and semi-arid regions in northwest China,one of the most sensitive areas for climate change,showing a trend of change from warm-dry conditions to warm-wet conditions (Shiet al.,2007;Yaoet al.,2015).Therefore,climate change has become a major driving force affecting the ecological and physiological processes of terrestrial forest ecosystems in northwest China,which affects the response stability and trends in the radial growth of trees (Liet al.,2015).We analyzed the climate response as reflected in the radial growth of Chinese pine trees at different altitudes in the eastern Qilian Mountains.Results show that drought was still a common limiting factor of radial growth of Chinese pine at the two altitudes,despite climate change from warm-dry to warm-wet conditions in the study region.As supporting evidence,the radial growth at the two altitudes show a significantly negative correlation with temperature in both September of the previous year and July of the current year (Figure 4a),and that at low altitude displayed a significantly positive correlation with total precipitation in July of the current year(Figure 4d).In addition,the radial growth of Chinese pine at the two altitudes significantly positively correlated with the drought index (PDSI) in July and August (Figures 4e and 4f).The correlation coefficients of first difference of chronology and climate factors also show the same results,which further corroborated that drought was a key factor restricting the radial growth of Chinese pine.
Drought is a major constraint of forest development and tree growth in arid and semiarid regions(Barberet al.,2000).In particular,our results also indicate that drought in the growing season became the common limiting factor of the climate for the two species in the last decades (Figure 4).Furthermore,evaporation in the study area is far greater than total rainfall.Thus,the effective soil moisture struggles to meet the needs of plants (Liet al.,2010).Since 1960,the mean annual temperature and total precipitation have been on the rise (Figure 2b).In order to eliminate the influence of dimension and variable itself and value,"Z-score standardization"was used to standardize data of annual mean temperature and total precipitation,and it was found that both temperature and precipitation show an upward trend.However,the increasing trend of temperature (0.428/10a) was faster than that of precipitation (0.202/10a).Therefore,mountain forest ecosystems in arid regions are particularly susceptible to drought stress and the radial growth of trees in those regions is considered to be especially sensitive to changes in drought (Zhanget al.,2016).First of all,the high temperature in the growing season may intensify the respiration and transpiration of trees,thus consuming the organic compounds fixed by trees with the warming of the climate.In addition,high temperatures may accelerate the evapotranspiration of soil water,which could reduce the available water of trees and lead to water deficits and growth restriction (Lavergneet al.,2015).Moreover,climate warming might also lead to a decrease in net accumulation of photosynthesis,an increase in respiration rate and a decrease in stomatal conductance,leading to a greater dependence of radial growth on drought stress (Wuet al.,2015;Lianget al.,2016).Secondly,the study areas are located inland of northwest China,far from the ocean,in an area with obvious temperate continental climate characteristics and belonging to a temperate semi-arid region.Although annual total precipitation increased in 1960-2018,the water-holding ability of the soil was still lower with less total precipitation and larger annual evaporation,steeper slopes and an open canopy (Table 1 and Figure 2a).Therefore,drought caused by high temperature and strong evaporation was an important factor affecting the radial growth of Chinese pine trees in this region(Liet al.,2010;Zhouet al.,2013).

Figure 6 Interannual variability of basal area increment(BAI)at different altitudes(the solid black line represents the 10-year moving average of BAI)
The radial growth of Chinese pine at the two altitudes in the study area was restricted by drought stress,but there were differences in the combination and strength of factors.The drought stress limiting trees in high-altitude is mainly caused by the single factor of higher temperature in the growing season,while that in low-altitude is mainly caused by the combination of lower precipitation and higher temperature in the growing season (Figure 2a).Many studies have also shown that the relationship between treering growth and climate is affected by altitude (Makinenet al.,2002;Linderholmet al.,2003;Tardifet al.,2003).The growth of Chinese pine trees at high altitude was negatively correlated with the temperature in September of the previous year and July of the current year.First of all,the high temperature at the end of the growing season of the previous year lengthens the growing period for trees.This would cause excessive consumption of the nutrients stored by the trees,and increases the risk of trees overwintering,thereby affecting the physiological activities of the trees in the second year (Jiaoet al.,2020).Secondly,the high temperature in the middle of the growing season could aggravate soil moisture evaporation,causing body moisture loss of trees and limiting physiological activities such as photosynthesis rate (Wanget al.,2003;Guoet al.,2004).
In addition,the growth of Chinese pine trees at low altitude was negatively correlated with the temperature in September of the previous year and July of the current year,and was positively correlated with precipitation in July of the current year (Figure 4d).This indicates that high temperatures may accelerate the transpiration rate of plants and increase the water demand,aggravating the evapotranspiration of soil water,reducing the available water content of soil and causing a lack of water in the root system,which is not conducive to the growth of trees (Takahashiet al.,2010;Zhanget al.,2010;Liet al.,2011).Studies on the climate response of the deciduous pine in the eastern Tianshan Mountains found that the growth of forest trees under the line was comprehensively affected by climate factors such as atmospheric temperature and precipitation,which also confirmed our research results (Houet al.,1999;Penget al.,2006).At the same time,low altitude areas had less precipitation,higher temperatures and intensified drought stress compared with high altitude areas,leading to increased sensitivity of radial growth to water availability (Hartl-Meieret al.,2014).In addition,the competition between species for resources gradually increases with decreased altitude,thus increasing the sensitivity of the radial growth of trees to dry drought conditions(Leonelliet al.,2002;Ettingeret al.,2011).
4.2 Evaluation of response stability to climate change of Chinese pine
As a valuable and widely used material,tree rings have the potential to provide millennial-scale climate measurement data,but the unstable response of radial growth to climate change is also a challenge in the application of traditional tree ring chronology (D'Arrigoet al.,2008).Our results show that the radial growth of Chinese pine at different altitudes had divergence response characteristics,with unstable nonlinear responses to climate change,showing that the temperature at the end of the growing season of the previous year increased the limiting effect on radial growth of trees at both altitudes,and drought in the middle of the growing season increased the limiting effect on radial growth of trees at low altitudes(Figure 5).
During recent decades,warmer climate conditions that were once favorable to radial growth seemed to become detrimental.A number of forests in Eurasia and western North America also exhibited shifts to negative correlations between summer temperature and radial growth,coupled with declining growth rates over the past few decades,with rising temperature-induced drought stress invoked as the dominant mechanism (Buermannet al.,2014).The shortage of available soil moisture might lead to the inhibition of root development,cambial activity,nutrient storage,foliage efficiency and the proportion of early wood(Andreu-Hayleset al.,2011;Xuet al.,2012).Climate warming is by now unequivocal,as is evident from observations of increases in global average temperatures (IPCC,2013).Climate warming poses the risk of much drier conditions,potentially leading to modified growth trends and response patterns of forests around the world,especially in drought-prone regions(Gazolet al.,2015).Since around the middle of the twentieth century,evidence of the reduced sensitivity of tree growth to temperature have been reported in species such asPicea glauca(Alaska),Picea abies(Alps),Picea rubens(southern Appalachian Mountains),andFagus sylvatica(the European Mediterranean Basin) (Lloyd and Fastie,2002;Büntgenet al.,2006;Tegelet al.,2014;Whiteet al.,2014).Similar results have also been recorded by several dendroclimatic studies on species such as Chinese pine (the Helan Mountains),Sabina przewalskii(the Qilian Mountains),Picea crassifolia(northeast Tibetan Plateau),andPinus koraiensis(the Changbai Mountains)in China(Zhanget al.,2009;Zhang and Wilmking,2010;Gaoet al.,2013;Yuet al.,2013).Combining these instances of unstable growth-temperature relationships,drought stress caused by warming might be a major constraint to the radial growth of Chinese pine in recent decades (D'Arrigoet al.,2008).Our results are also in good agreement with these prior studies.Therefore,we believe that the intensification of drought stress might be a primary reason for the "divergence problem" in the eastern Qilian Mountains.
4.3 Potential influence of climate change on the radial growth trends of Chinese pine at different altitudes
Global average temperature rises by about 0.02 ℃/a under the medium emission scenario and about 0.04 ℃/a under the high emission scenario(IPCC,2013).There are more striking upward trends of temperature in the Qilian Mountains in the future based on the GSFLOW model with an increase of 0.03 ℃/a under the medium emission scenario and 0.06 ℃/a under the high emission scenario (Wuet al.,2019).Therefore,this shows that the intensity of climate warming in our study region exceeds the global average.The growth of Chinese pine shows the same trend at both elevation gradients,according to the 10-year moving average of BAI (Figures 6a and 6b).The growth of Chinese pine at high altitude increased before 1968 and then decreased sharply,and that at low altitude show a continuous upward trend before 1993 and then a slow downward trend.At the same time,the mean temperature was stable before 1996 and rose rapidly from 1996 to 2018.This conclusion is consistent with previous results of the growth trends of trees and growth-climate relationships.
Although plants can gradually adjust their strategies to adapt to climate change,the growth response of trees is likely to be limited by a range of physiological thresholds (Dayet al.,2002).Within the threshold range,tree growth was positively correlated with temperature,but negatively correlated with temperature beyond a certain threshold.The temperature has risen significantly since the 1990s(Figure 2c),and the variation trends of BAI at the upper and lower limits of the altitude distribution of Chinese pine were obviously different after the temperature increase,indicating that trees at the upper and lower limits of the altitude distribution of Chinese pine have different modes of responding to climate warming.Moving correlation analysis shows that the temperature rise significantly limited the radial growth of Chinese pine,demonstrating that the negative response of Chinese pine to temperature was gradually strengthened with time (Figure 5).This is consistent with previous research results,showing that the radial growth of Chinese pine,Juniperus przewalskiiand Qinghai spruce in the eastern Qilian Mountains was limited by drought with the rapid rise of temperature (Zhanget al.,2011;Zhanet al.,2019).If low altitude temperatures continue to rise,especially in the growing season,this may lead to further weakening of the radial growth and increased mortality of the Chinese pine,thus influencing the distribution of low elevation Chinese pine trees.As the altitude rises with increased precipitation and decreased temperature,Chinese pine at high altitude show a significant "growth divergence phenomenon" in 1968 (Figure 6a).The radial growth shows a negative temperature-dependent response,similar to that observed at low altitude,based on the correlation analysis.The reasons for Chinese pine growth at high altitude also being limited by drought stress might be related to factors such as relatively open canopy at high altitude,steep slopes,and high solar radiation,which lead to high soil evapotranspiration loss and low water holding capacity.In addition,trees at high altitudes are more sensitive to climate change,showing an earlier downward trend than those at low altitudes (Figure 6b).The main reason was that trees at high altitude had been restricted by low temperature and were less adaptable to drought restrictions after climate change.
The total precipitation in the study areas increased significantly from 1960 to 2018 (Figure 2b).However,the increase in annual precipitation did not increase the growth rate of trees correspondingly.It may be that drought stress caused by heat was not alleviated by increased precipitation due to rapid temperature rise (Figure 4c).As climate warming intensifies,forest ecosystem has been seriously affected in the southeastern France Mediterranean Mountains,interior Alaska,northern Canada,and boreal North America,Mongolia,Qinghai-Tibet Plateau,Qilian Mountains in Asia(D'Arrigoet al.,2009;Lebourgeoiset al.,2012).If temperatures continue to rise in the future,we predict that the structure and function of forests will change and the mortality and degradation of trees will increase in arid and semi-arid areas (Liuet al.,2013).
5 Conclusions
Chinese pine at different altitudes in the Qilian Mountains show differing temporal response stability and radial growth patterns under changing climate conditions since the 1960s.Based on the results,drought caused by high temperature was identified as a common limiting factor for the radial growth of Chinese pine at the two altitudes,although the climate changed from warm-dry to warm-wet conditions in the study areas.Furthermore,the effects of drought stress on tree growth gradually increased from high to low altitudes.Therefore,the response of tree species to climate provides useful information for understanding the adaptation strategies and ecological thresholds of forest ecosystems.The effects of climate change on forest ecosystems can be minimized by establishing corresponding forest management measures based on the differences in response characteristics between tree growth and climate at different altitudes.In addition,the radial growth of Chinese pine at different altitudes in response to the main limiting climatic factors was unstable and presented a synchronous radial growth pattern,increasing at first and then decreasing.Therefore,the evaluation of response stability of radial growth to climate change should be considered in light of historical climate reconstruction and simulation of forest development trends.Further strengthening of forest protection and water resource management is necessary to cope with further climate warming in the future,especially for mountain forest ecosystems in arid and semi-arid regions.
Acknowledgments:
This research was supported by the National Natural Science Foundation of China (Grant No.41861006),Project/ Supported by State Key Laboratory of Earth Surface Processes and Resource Ecology (Grant No.2020-KF04) and the Research Ability Promotion Program for Young Teachers of Northwest Normal University (NWNU-LKQN2019-4).We also thank anonymous referees for helpful comments on the manuscript.
 Sciences in Cold and Arid Regions2021年6期
Sciences in Cold and Arid Regions2021年6期
- Sciences in Cold and Arid Regions的其它文章
- Cryogenic wedges on the NE Qinghai-Tibet and Ordos Plateaus:Their characteristics,origin and OSL dating
- High-precision measurements of the inter-annual evolution for Urumqi Glacier No.1 in eastern Tien Shan,China
- Editor-in-Chief Yuanming Lai
- Satellite-measured water vapor isotopologues across the Tianshan Mountains,central Asia
- Long-term effects of gravel-sand mulch thickness on soil microbes and enzyme activities in semi-arid Loess Plateau,Northwest China
- Seasonal variation of airborne fungi of the Tiantishan Grottoes and Western Xia Museum,Wuwei,China