
哺乳仔猪断奶前后肠道微生物培养组学研究
2021-02-06 05:47王志林魏文康井晓欢鄢秋龙蒋宗勇刘文华胡译尹
动物营养学报 2021年1期
董 博 王志林 魏文康,3 井晓欢 刘 石 鄢秋龙 蒋宗勇 刘文华 胡译尹 陈 庄*
(1.广东省农业科学院农业生物基因研究中心,广东省农作物种质资源保存与利用重点实验室,广州510640;2.广东省畜禽疫病防治研究重点实验室,农业农村部兽用药物与诊断技术广东科学观测实验站,广州510640;3.岭南现代农业科学与技术广东省实验室茂名分中心,茂名525000;4.深圳国家基因库,深圳510083;5.大连医科大学微生物教研室,大连116044;6.广东省农业科学院,广州510640)
肠道是营养物质消化吸收、微生物与宿主互作的主要部位。肠道微生物作为肠道生态系统的重要部分和调节因子,其健康状态关系着断奶仔猪的正常生长和发育[1]。仔猪断奶应激一直是困扰养猪业的难题,断奶前后仔猪胃肠道微生物区系也发生剧烈的变化。断奶前后仔猪胃肠道微生物区系的基因组学研究和功能活性的研究已有报道[2-16]。Li等[2]研究了断奶前后仔猪结肠内容物的微生物基因组,发现分别含有1 584和1 157个操作分类单元(operational taxonomic unit,OTU),其中分别有137和85个特定OTU。尽管如此,目前人和动物肠道微生物组研究中,仍然有超过一半宏基因组测序不能得到明确的菌种信息,更不能获得分离纯化的单菌株,以进行表型、功能以及与宿主的互作研究[17-22],因此需要对传统的菌株筛选方法进行优化改造。培养组学(culturomics)是一种使用多种培养条件和16S rRNA测序相结合,对样品中的微生物进行系统地筛选分离培养的方法[17,23-32]。培养组学方法可以将之前认为不可培养的菌株培养起来,较大程度地发现潜在的新菌株,丰富现有的可培养肠道菌株资源库,为开发缓解断奶应激的益生菌打下基础。在畜禽肠道微生物培养组学研究方面,2017年,Ferrario等[33]首次利用培养组学方法对百慕大野鸡、蛋鸡和肉鸡盲肠内容物进行了培养组研究,获得并鉴定了417株菌株(包括4个门、15个属和43个种),并对相应的培养样本进行了宏基因组分析;2018年,Medvecky等[34]培养了鸡盲肠内容物,共获得了7个门、133株不同的厌氧菌种,并全部进行了全基因组测序分析,为开发益生菌提供了较全面的菌株库和基因资源;2020年,德国马普所建立并公开了猪的肠道细菌库,包括9个门、39个科和112个种,对38个新的类群进行了分类学描述,包括21个新种和17个新属,并进行了扩增子和宏基因组测序,以及功能注释、抗菌分析和益生菌开发的详细研究[35],上述研究工作还在继续。目前国内应用培养组学方法在猪肠道微生物全面筛选培养方面和益生机制的研究才刚刚开始,本研究使用培养组学方法,对仔猪断奶前后肠道微生物进行全面筛选、培养和鉴定分析。
1 材料与方法
1.1 断奶前后仔猪肠道内容物样本
选择断奶前后的丹系“杜长大”三元杂仔猪各3头(雌性,健康状况良好,同一窝仔猪),分别于18日龄(断奶前,体重2.8 kg)和60日龄(断奶后,体重35 kg)屠宰。无菌收集仔猪回肠和结肠内容物,并立即使用培养组学方法,对样本中的细菌进行筛选培养。仔猪断奶前后均未用抗生素。
1.2 细菌培养方法
选择25种培养基,包含5%羊血-104、5%羊血-脑心浸液(brain heart infusion,BHI)、5%羊血-DM、5%羊血-GMM、5%羊血-R2A、5%羊血-SCH、5%羊血-孢子、5%羊血-哥伦比亚、Kana-104、Kana-BHI、Kana-哥伦比亚、常规-104、常规-2216、常规-27、常规-98、常规-AM、常规-BHI、常规-DM、常规-GMM、常规-GYM、常规-NA、常规-R2A、常规-SCH、常规-孢子和常规-哥伦比亚,分别对样品进行稀释培养。
1.2.2 培养条件
对每份样品分别使用厌氧(使用Bactron Ⅳ-2厌氧培养箱,Shellab)和好氧[使用KB240(E5.1)低温培养箱,Binder]条件进行37 ℃培养。
1.2.3 试验过程
样品处理:好氧处理在超净工作台中,厌氧处理在厌氧培养箱中进行。每份样品分别进行好氧和厌氧处理。处理步骤如下:
常规培养:1)每份样品取样各1 g,置于含有9 mL pH 7.2磷酸盐缓冲液(PBS)的无菌15 mL离心管中混匀;2)使用pH 7.2的PBS对样品进行10倍系列稀释;3)每份样品系列稀释后为10-1~10-8,每个稀释梯度各10 mL;4)选择10-5、10-6和10-7稀释梯度,各吸取100 μL稀释液,在相应的琼脂培养基平板上进行涂布;5)在37 ℃下分别进行厌氧和好氧静止培养,培养48 h后,进行菌株挑取。
富集培养:1)每份样品取样各1 g,置于15 mL无菌离心管中,并加入相应液体培养基9 mL,振荡混匀;2)分别置于厌氧、好氧环境下37 ℃进行培养;3)分别于第1、3、7、10、15、21和30天取1 mL培养物,使用相应液体培养基进行10-1~10-8系列稀释;4)取10-5、10-6和10-7稀释梯度各100 μL涂布于对应固体培养基上;5)于恒温摇床中37 ℃和180 r/min进行厌氧和好氧培养,培养48 h后,进行菌株挑取。
1.3 16S rRNA全长测序
应用27F和1492R引物分别对每种菌落的基因组DNA进行PCR扩增并进行Sanger测序,获得约1.5 kb的16S rRNA基因全长序列,并比对到微生物数据库(NCBI数据库和古菌16S rRNA标准数据库),相似度>98.65%为确定菌株的种属,相似度≤98.65%则认为此菌株为疑似新种。整个培养组学工作流程见图1。
第三代图书馆服务平台将更加贴近读者的生活、阅读需求及知识获取和利用方式,为读者提供高度定制化的服务。通过对读者互联网社交媒体、多种文化生活场所的集成与覆盖,对读者兴趣与阅读行为的深入分析,从资源组织与内容服务等多个角度实现对读者服务的高度定制化。
2 结果与分析
2.1 培养方法和菌株挑取
本研究基于培养组学方法,使用25种培养基对样品中的细菌进行筛选培养,共获得1 385株细菌。通过观察涂布平板上的菌落形态的差异挑取菌株,其中常规-R2A、常规-哥伦比亚和常规-104这3种培养基所筛选挑取的菌株数目最多,分别为131、114和113株;另外,孢子、DM、BHI、GMM、SCH这4种培养基上菌株也生长情况良好;常规-GYM、常规-27、常规-2216和常规-AM培养基培养的菌株数目较少(表1)。

a:仔猪断奶前后回肠和结肠内容物 ileum and colon contents of piglets before and after weaning;b:样品先在PBS中稀释 samples were first diluted in PBS;c:在25种液体培养基采用无氧和有氧条件37 ℃进行培养 incubate diluted samples in 25 kinds of liquid medium under anaerobic and aerobic conditions at 37 ℃;d:平板筛选的菌落甘油保存并提取基因组DNA用于鉴定 preserve the colonies selected by the corresponding plate in glycerol stock and extract the genomic DNA for identification;e:16S rRNA基因测序分析及菌种鉴定 16S rRNA gene sequencing analysis and strain identification。

表1 每种培养基培养的菌株数目
实际培养过程中,根据猪肠道的特点,在培养基中会添加一些生长因子,如微量元素、维生素和脱纤维羊血等[36],尽可能将样品中的细菌在实验室培养获得。同时,涂布平板培养时间也较重要,大部分培养获得的菌株,都是在24~48 h中获得[26],但是较难培养获得的菌株一般培养时长超过48 h[37-38]。对于使用常规培养方法较难培养的拟杆菌和梭菌属,其中拟杆菌本研究使用常规-SCH和Kana-104培养基培养出来了,培养出的拟杆菌均来自仔猪断奶后结肠样本;梭杆菌本研究使用5%羊血-R2A、5%羊血-哥伦比亚、5%羊血-104、5%羊血-R2A、常规-R2A和常规-BHI这6种培养基培养出来了,这6种培养基中4种都添加了5%羊血,显示梭杆菌比较容易用5%的脱纤维羊血培养出来,培养出的这9株梭杆菌除了1株来自回肠,其余8株都来自结肠样本,显示梭杆菌可能和结肠中降解纤维多糖方面的功能有关[39]。
2.2 菌株鉴定
2.2.1 不同培养条件及门、属、种水平的细菌鉴定
由表2和表3可知,筛选培养鉴定的1 385株菌株,分布于5个门、29个属和86个种。仔猪断奶前回肠细菌门水平有厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)和梭杆菌门(Fusobacteria)3个门;断奶后回肠有厚壁菌门和变形菌门这2个门,此2个门也存在断奶前回肠。仔猪断奶前结肠菌株分布在厚壁菌门、拟杆菌门(Bacterioidetes)、变形菌门、放线菌门(Actinobacteria)和梭杆菌门;断奶后结肠菌株分布在厚壁菌门、拟杆菌门、变形菌门和放线菌门,断奶后结肠没有培养出梭杆菌门。仔猪断奶前回肠培养了305株菌,结肠培养了386株菌;断奶后回肠培养了220株菌,结肠培养了474株菌(图2)。
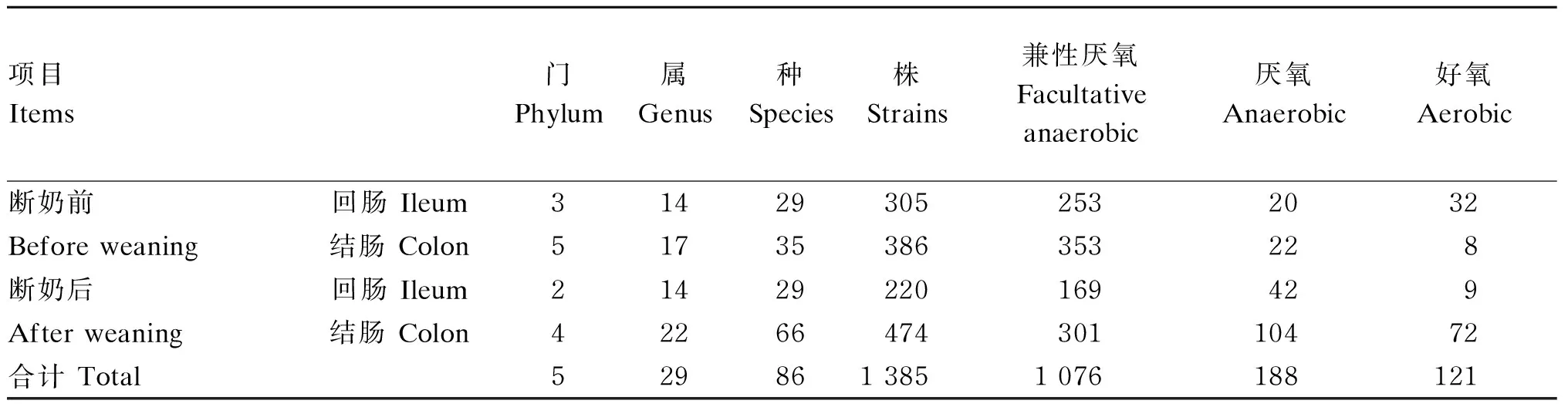
表2 仔猪断奶前后回肠和结肠鉴定细菌门、属、种分布及需氧厌氧特性

续表3项目 Items组成 Composition(Corynebacterium xerosis)、6株铅黄肠球菌(Enterococcus casseliflavus)、17株粪肠球菌(Enterococcus faecalis)、11株海氏肠球菌(Enterococcus hirae)、8株泰国肠球菌(Enterococcus thailandicus)、1株艾伯特埃希菌(Escherichia albertii)、43株大肠杆菌(Escherichia coli)、125株费格森埃希菌(Escherichia fergusonii)、10株旱獭埃希菌(Escherichia marmotae)、1株Faecalicatena orotica、10株死亡梭杆菌(Fusobacterium mortiferum)、1株产气克莱伯氏菌(Klebsiella aerogenes)、1株肺炎克莱伯氏菌(Klebsiella pneumoniae)、2株嗜酸乳杆菌(Lactobacillus acidophilus)、2株敏捷乳杆菌(Lactobacillus agilis)、73株淀粉乳杆菌(Lactobacillus amylovorus)、11株Lactobacillus coleohominis、9株卷曲乳杆菌(Lactobacillus crispatus)、11株发酵乳杆菌(Lactobacillus fermentum)、2株格氏乳杆菌(Lactobacillus gasseri)、60株约氏乳杆菌(Lactobacillus johnsonii)、3株乳酸乳杆菌(Lactobacillus lactis)、36株黏膜乳杆菌(Lactobacillus mucosae)、242株罗伊氏乳杆菌(Lactobacillus reuteri)、16株唾液乳杆菌(Lactobacillus salivarius)、149株阴道乳杆菌(Lactobacillus vaginalis)、5株埃氏巨型球菌(Megasphaera elsdenii)、1株Novosphingobium aromaticivorans、1株Paenibacillus polysaccharolyticus、1株索氏梭菌(Paeniclostridium sordellii)、1株狄氏副拟杆菌(Parabacteroides distasonis)、25株Pelistega suis、2株布氏假单胞菌(Pseudomonas brenneri)、2株荧光假单胞菌(Pseudomonas fluorescens)、1株边缘假单胞菌(Pseudomonas marginalis)、31株草假单胞菌(Pseudomonas poae)、2株平凡假单胞菌(Pseudomonas trivialis)、1株维罗纳假单胞菌(Pseudomonas veronii)、2株鲍氏志贺氏菌(Shigella boydii)、68株弗氏志贺氏菌(Shigella flexneri)、23株宋内志贺氏菌(Shigella sonnei)、1株森林土源芽孢杆菌(Solibacillus silvestris)、2株松鼠葡萄球菌(Staphylococcus sciuri)、1株人葡萄球菌(Staphylococcus hominis)、1株腐生葡萄球菌(Staphylococcus saprophyticus)、7株嗜根窄食单胞菌(Stenotrophomonas rhizophila)、13株非解乳糖链球菌(Streptococcus alactolyticus)、35株猪肠链球菌(Streptococcus hyointestinalis)、10株猪阴道链球菌(Streptococcus hyovaginalis)、1株副血链球菌(Streptococcus parasanguinis)、4株鲁布纳链球菌(Streptococcus rubneri)、22株豚鼠韦荣氏球菌(Veillonella caviae)、34株麦格纳韦荣氏球菌(Veillonella magna)和3株食窦魏斯氏菌(Weissella cibaria)
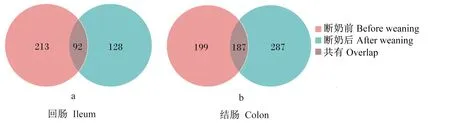
图2 仔猪断奶前后回肠和结肠筛选菌株韦恩图比较
2.2.2 仔猪断奶前后回肠和结肠细菌属种水平比较
由表2可知,在属种水平,仔猪断奶前回肠分布有14个属、29个种和305株的细菌,断奶后回肠分布有14个属、29个种和220株的细菌;断奶前结肠分布有17个属、35个种和386株的细菌,断奶后结肠分布有22个属、66个种和474株的细菌。根据表2总结的细菌的好氧厌氧特性来看,厌氧和兼性厌氧菌为1 264株,好氧菌为121株。仔猪结肠的厌氧菌数目明显多于回肠,结肠的厌氧菌有126株、回肠有62株,这与之前肠道氧含量的结论[6,9-10]是一致的。另外,仔猪断奶后回肠和结肠的厌氧菌和兼性厌氧菌数目都有很大程度的增加,显示仔猪断奶后肠道菌群的多样性增加,厌氧菌所占菌群的比例也有很大程度的增加。此外,仔猪断奶前后回肠有92株菌是共有的,结肠有187株菌是共有的,结肠共有的比例大于回肠,这表明回肠菌群结构变化大于结肠(图2)。
虽然培养组学方法得到的菌株数目只能粗略地反映菌种的丰度,但从图3和图4可以看出,与断奶前相比,仔猪断奶后回肠中乳杆菌属(Lactobacillus)菌株数目有很大程度的减少,这与断奶前仔猪主要食源为母乳,而乳杆菌具有促进母乳脂肪消化和吸收的功能是一致的。另外,仔猪断奶前回肠、结肠中的潜在致病菌,如韦荣氏球菌属(Veillonella)、链球菌属(Streptococcus)等菌株数目都很高,可能是母乳的抗体有较强的保护作用,而在断奶后42 d,肠道菌群初步成熟,回肠中这些条件致病菌株数目就有很大程度的减少。但是,仔猪断奶后结肠中蛭弧菌属(Bdellovibrio)、短杆菌属(Brevibacterium)、纤维微菌属(Cellulosimicrobium)、艰难梭菌属(Clostridioides)、窄食单胞菌属(Stenotrophomonas)、魏斯氏菌属(Weissella)、新鞘脂菌属(Novosphingobium)、类芽孢杆菌属(Paenibacillus)、假单胞菌属(Pseudomonas)、土壤芽孢杆菌属(Solibacillus)及葡萄球菌属(Staphy-lococcus)这些菌属菌株在断奶前都不存在,总体上断奶后结肠菌属多样性要高很多,并且断奶后结肠中韦荣氏球菌属、梭杆菌属(Fusobacterium)、拟杆菌属(Bacteroides)和巨型球菌属(Megasphaera)这几个属的菌株数目有明显的减少,而断奶后结肠中埃希氏杆菌属(Escherichia)、志贺氏菌属(Shigella)、芽孢杆菌属(Bacillus)、纤维微菌属、艰难梭菌属和假单胞菌属这6个属菌株数目明显增多,芽孢杆菌属和纤维微菌属是和纤维素降解有关的菌,埃希氏杆菌属和志贺氏菌属也是大肠中常见的菌属,也是一种条件致病菌。
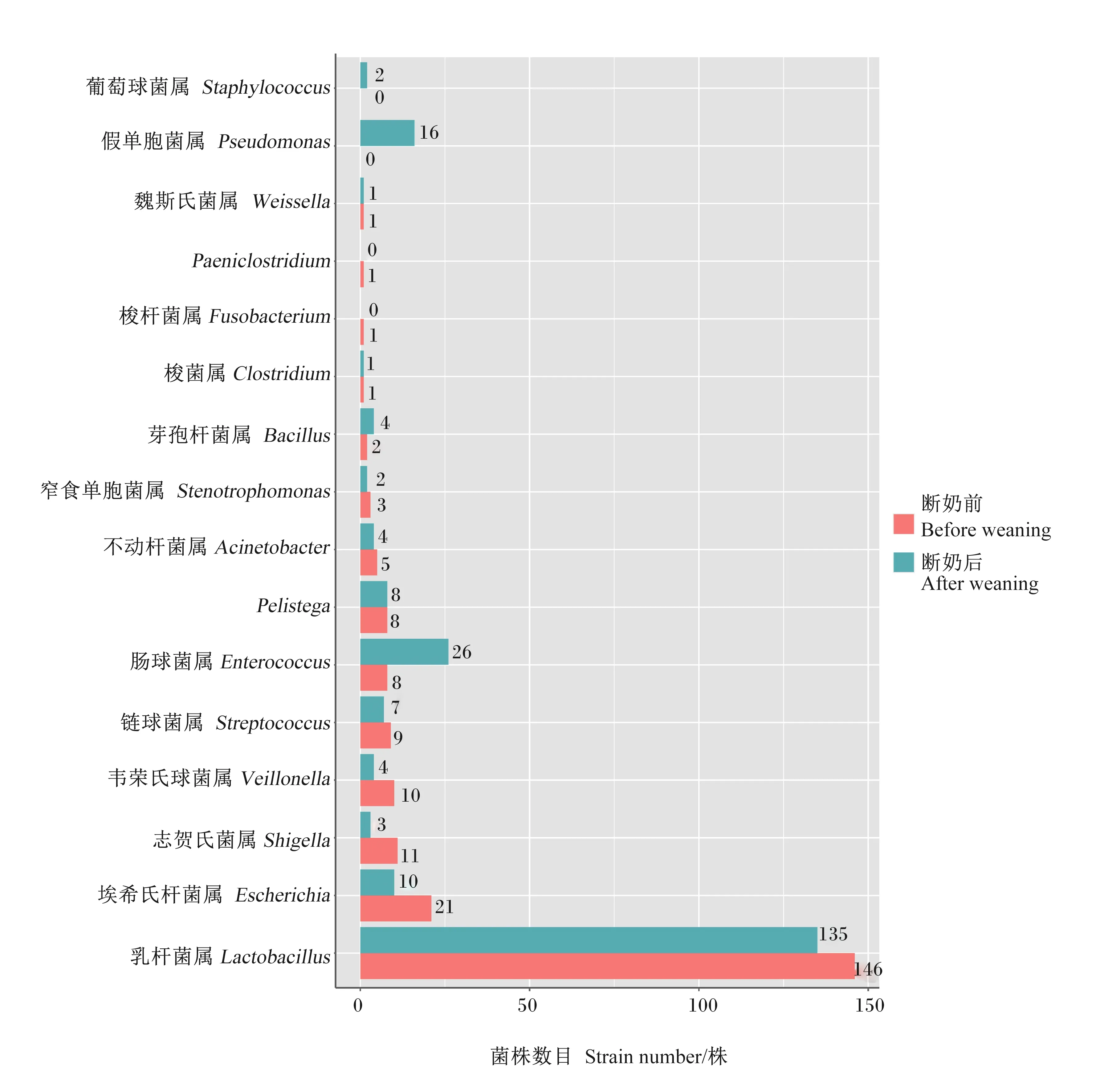
图3 仔猪断奶前后回肠培养的菌株在属水平上的分布柱状图
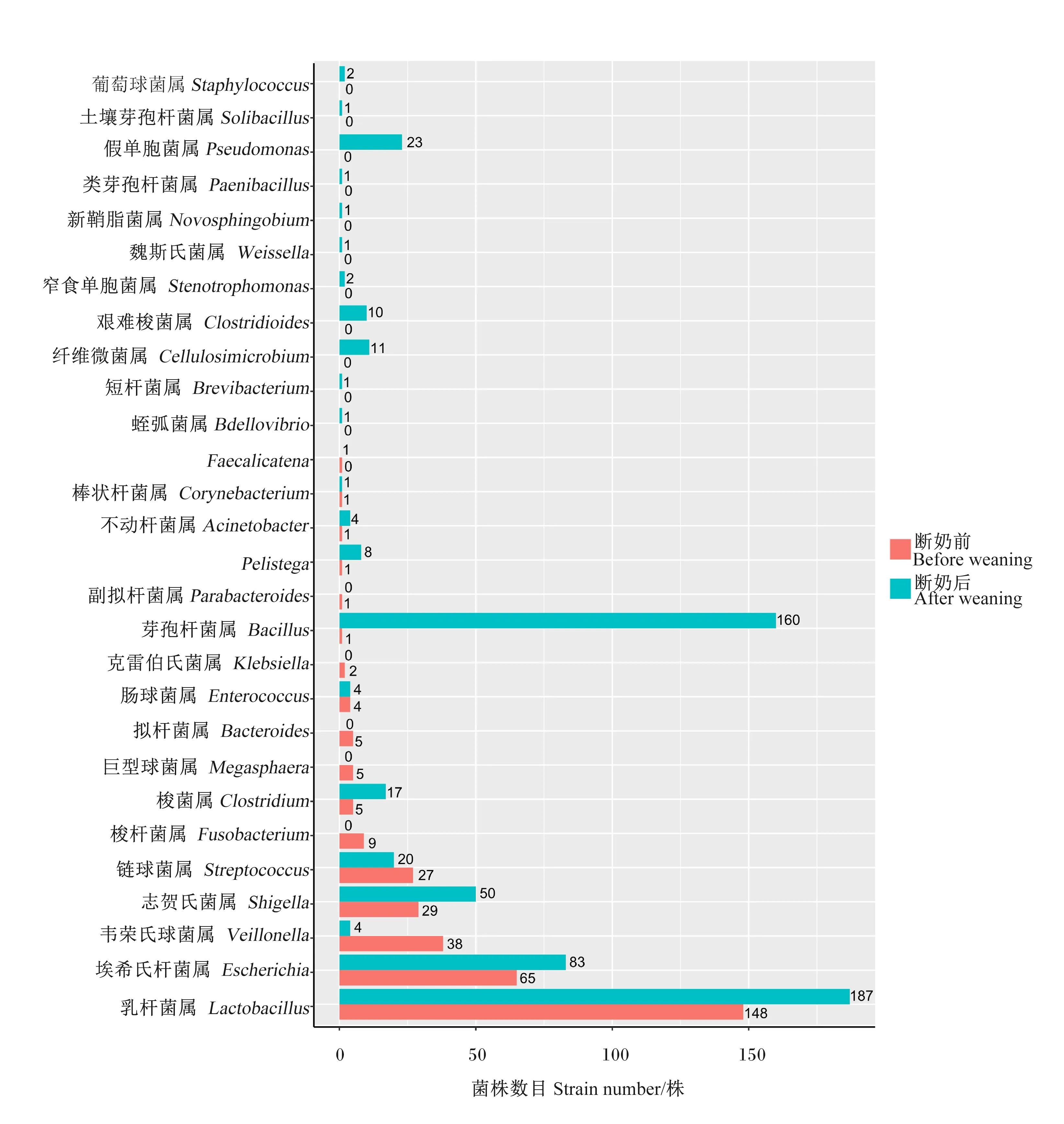
图4 仔猪断奶前后结肠培养的菌株在属水平上的分布柱状图
从图5可以看出,仔猪断奶前后回肠菌种数目都为29种,但是从属水平看,断奶后回肠中不动杆菌属(Acinetobacter)、梭菌属(Clostridium)、埃希氏杆菌属、梭杆菌属、Pelistega、志贺氏菌属、窄食单胞菌属、链球菌属、韦荣氏球菌属和魏斯氏菌属种数目都下降到0,而断奶后回肠中芽孢杆菌属、肠球菌属(Enterococcus)、乳杆菌属、假单胞菌属、葡萄球菌属种数目都有所增加。从图6可以看出,与断奶前相比,仔猪断奶后结肠中菌种数目明显增多,增加的包括梭菌属、芽孢杆菌属、肠球菌属、短杆菌属、纤维微菌属、艰难梭菌属和假单胞菌属等,而乳杆菌属等少数属种数目则有所减少。
2.2.3 仔猪断奶前后肠道微生物培养组学发现的疑似新菌种
根据16S rRNA全长与NCBI微生物数据库结果比对,相似度≤98.65%即为潜在新菌种[40],本研究的培养方法一共获得116株疑似新种(表4),其中乳杆菌属最多共53株,其次是假单胞菌属共18株,平凡假单胞菌(Pseudomonastrivialis)和屎肠球菌(Enterococcusfaecium)各8株,费格森埃希菌(Escherichiafergusonii)5株,Faecalicatenaorotica、无害梭菌(Clostridiuminnocuum)、丁酸梭菌(Clostridiumbutyricum)、糖丁酸梭菌(Clostridiumsaccharobutylicum)、黏膜乳杆菌(Lactobacillusmucosae)、埃氏巨型球菌(Megasphaeraelsdenii)、狄氏副拟杆菌(Parabacteroidesdistasonis)、猪肠链球菌(Streptococcushyointestinalis)和鲁布纳链球菌(Streptococcusrubneri)等各1株。
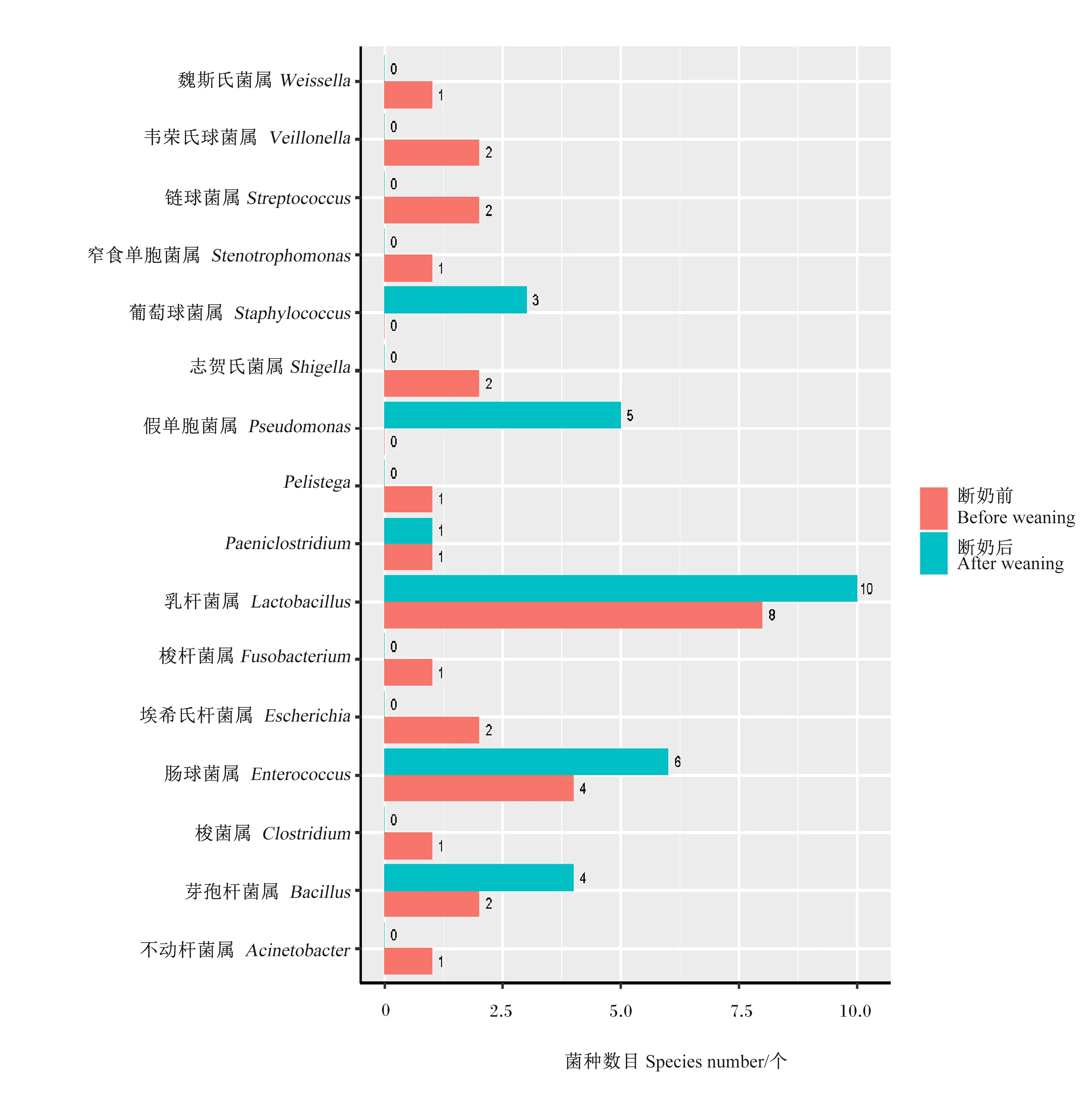
图5 仔猪断奶前后回肠培养的菌种在属水平上的分布柱状图
3 讨 论
近年来,培养组学方法在获得潜在新菌种、丰富可培养菌株资源库及菌株的益生机制研究方面,已显现出巨大的优势,并已在人体肠道微生物和相关医学研究中成为新的热点。在畜牧业研究领域,培养组学也已在菌株与宿主相关作用、病原微生物及益生机制和动物营养学方面的研究中显现出其潜在的优势,但是有关研究在国内却鲜见报道。
3.1 培养组学条件和菌株筛选鉴定
为了尽可能全面地培养肠菌,本研究对仔猪断奶前后的回肠和结肠内容物组合使用25种培养基和培养条件(包括厌氧、纤维羊血和营养因子),还针对肠道菌株筛选的特点,如多会产生孢子的情况,添加使孢子萌发的成分,在培养中对样品进行了一些预处理、预培养和富集等,共培养获得1 385株细菌,与之前的宏基因组研究报道的1 157~1 584个OTU[2]比较相对较少,一些宏基因注释出来的丰度较低的门[如螺旋体门(Spirochaetes)和软壁菌门(Tenericutes)等]和属[如罗斯氏菌属(Roseburia)、多雷亚菌属(Dorea)、Blautia、普雷沃氏菌属(Prevotella)等]没有培养出来[41],其可能原因是挑菌过程中为了避免过多重复并没有全面挑取,或者一些菌种需要更长的平板培养时间,如螺杆菌属(Helicobacter)的一些菌种需要几天的时间[37],一些分支杆菌属(Mycobacterium)的菌种需要几周的时间[38]。因此需要对培养组学方法进一步研究开发,以获得更全面的猪肠道微生物菌库[24]。
3.2 仔猪断奶前后回肠和结肠微生物培养组学和测序结果比较
在门水平上,仔猪断奶前后回肠和结肠可培养的菌株分类变化不大,主要分布在厚壁菌门、拟杆菌门、变形菌门、放线菌门和梭杆菌门这5个门,其中厚壁菌门的相对丰度和多样性最高,这些和之前的宏基因组测序的研究结果[6]是一致的。宏基因组注释出来一些丰度较高的属如乳杆菌属、巨型球菌属、芽孢杆菌属、拟杆菌属和埃希氏杆菌属等,培养的结果中也有,而培养结果中的新鞘脂菌属、类芽孢杆菌属、Paeniclostridium、Pelistega、土壤芽孢杆菌属和窄食单胞菌属这些属未见于一些测序结果中[8,39,41-44]。可见培养组学和测序的结果是互相印证和补充的关系[31]。测序研究显示,对仔猪断奶后利用饲粮纤维比较重要的肠道菌属梭菌属、肠球菌属、葡萄球菌属[39]在本研究中也培养出来,这些菌可以作为仔猪适应断奶应激的候选益生菌。在种水平上,仔猪断奶前后回肠菌种数目未有变化,而断奶后结肠菌种数目明显增多。

表4 仔猪断奶前后肠道微生物培养组学发现的疑似新菌种(相似度≤98.65%)

续表4最相似属 Most similar genus最相似种 Most similar species菌株数目 Strain number/株链球菌属 Streptococcus非解乳糖链球菌(Streptococcus alactolyticus)2猪肠链球菌(Streptococcus hyointestinalis)1鲁布纳链球菌(Streptococcus rubneri)1韦荣氏球菌属 Veillonella麦格纳韦荣氏球菌(Veillonella magna)1合计 Total116
本研究结果表明,与断奶前相比,仔猪断奶后回肠中乳杆菌丰度发生了明显下降。仔猪肠道大部分菌群最开始是从母乳中获得并定植于肠道[45],母乳中乳酸菌比例较大,所以断奶前乳酸菌有优势,断奶前拟杆菌属、埃希氏杆菌属和梭杆菌属也比较丰富,这与宏基因组测定断奶前后菌属的变化结果相一致[8,12,44]。仔猪断奶时会突然转换到含有谷物和较高粗蛋白质的固体饲料[42,45],在这个转换期间,宏基因组分析显示回肠中作为早期病原菌拮抗的主要参与者之一乳杆菌属的数量突然减少[6],而结肠中梭菌属和包含大肠杆菌(Escherichiacoli)等变形菌门在内的有害菌在断奶后都有增加,拟杆菌属则有所减少[12],这与本研究的仔猪断奶前后结肠培养结果是一致的,这些菌属的变化大大增加患胃肠疾病的风险。断奶后,肠道菌群逐渐成熟,拟杆菌门丰度又逐渐上升[44];培养结果显示断奶应激降低了仔猪回肠种属多样性[42],而断奶后结肠菌种多样性的提高是由于可被结肠微生物代谢的纤维和多糖类饲粮的添加,这与宏基因组α多样性研究的结果是一致的;刚断奶的仔猪肠道菌属多样性先降低,然后从断奶后期到长成期又会上升,并超过刚断奶期[39,43]。
本研究中,仔猪回肠和结肠培养的菌群在门水平有很大不同,这与宏基因组数据结果一致,回肠培养出的主要菌群是厚壁菌门,占95%以上[6],而结肠中厚壁菌门、拟杆菌门和变形菌门都有一定比重,其中厚壁菌门和拟杆菌门总共占比超过90%,这和宏基因组门水平变化趋势研究结果[41]是一致。这种变化趋势从培养组学结果来看回肠中存在厌氧杆菌属(Anaerobacter)和Turicibacter较多,而结肠中存在普雷沃氏菌属、颤杆菌属(Oscillibacter)和琥珀酸弧菌属(Succinivibrio)较多[9]。宏基因组结果显示,在属水平上,断奶后拟杆菌属让位于梭菌属[13,45],这与本研究结果一致。结肠比回肠的含氧量要低,pH更高,适合多种厌氧和兼性厌氧菌生存。本研究中,虽然肠道菌群总体上厌氧菌占优势,但是结肠的厌氧菌和兼性厌氧菌的总和要明显高于回肠相应的比例[9,41]。
3.3 培养出来的潜在新菌种
本研究中,共发现了116株疑似新菌,更多菌株是第1次在猪肠道中发现,可以丰富有潜在应用价值细菌的猪肠道细菌库,有效拓展对仔猪肠道菌群多样性和仔猪肠道菌群及营养代谢关系的认识,实现对潜在益生菌更全面有效地捕获。
不同猪品系遗传因素对肠道菌群种属水平的影响较大,对15周龄的纯种猪品系如“杜洛克”、“长白”和“约克夏”品系之间的比较显示,这3种品系的肠道菌群在门水平上具有较高的一致性,均为拟杆菌门和厚壁菌门占主要,但是在属水平上,链型杆菌属(Catenibacterium)、考拉杆菌属(Phascolarctobacterium)和Subdoligranulum在“杜洛克”品系中更丰富,而小杆菌属(Dialister)在“约克夏”品系中更丰富[13]。因此,以后可以开展对中国本地猪的肠道培养研究,丰富猪肠道菌种库的遗传背景异质性,获取更全面的新型肠道细菌菌株资源。本研究证明,通过培养组学分离出的很多仔猪肠道菌群的多样性和丰度是迄今未知,这扩大了可培养的仔猪肠道微生物资源。
综上所述,只有通过各种培养方法,将样品中的微生物在实验室全面地筛选获得,并对其进行全基因组测序分析及功能注释,才能从根本上丰富肠道微生物菌种资源库和功能基因库,扩展对肠道菌群的全面认识,为仔猪营养研究和饲料益生菌制剂开发打下基础[23,28]。
本研究采用培养组学方法研究健康仔猪肠道微生物谱,其研究已深入到菌种水平,优于现有的基于二代测序技术的免培养分析方法。通过培养组学技术,我们能够发现潜在的新菌种,后续也能够对这些菌种进行全基因组测序分析及益生菌开发。并能够对获得的猪源益生菌进行益生机制深入研究。益生菌具有物种特异性,相比其他来源的益生菌,猪源性益生菌开发为微生态制剂,应用在猪营养调控上,将增加其定植概率和功效发挥[44,46],帮助在无抗养殖背景下解决仔猪断奶应激问题。
因此,在目前猪肠道基因集已经构建的基础上,展望未来的猪肠道微生物组学和营养研究是多组学、纯培养和生态特征的整合分析[44,47]。未来会针对培养的菌种库,基于比较基因组的方法,以菌种或亚种为单位研究不同菌种的遗传多样性,比较风险因子、功能代谢特性和菌株互补情况,展示常见益生菌的基因组特性及与生态位的关系,为解决仔猪断奶应激的益生菌的选择和复配提供参考。
4 结 论
本研究使用培养组学方法,对仔猪断奶前后肠道微生物进行全面筛选培养,共筛选获得1 385株厌氧、好氧及兼性厌氧菌株,共计获得5个门、29个属和86个种菌,其中包含梭杆菌、拟杆菌等在以前的研究中较难分离获得的菌株,以及一些如乳酸菌等具有潜在应用价值的益生菌,更为重要的是其中包含116株疑似新菌种。
致谢:
感谢深圳国家基因库的赵姣博士、广东省农业科学院动物卫生研究所彭新宇研究员、深圳倍森生物科技有限公司李胜辉工程师和中山大学生物技术系黄昱祈对文稿所提的宝贵意见。
猜你喜欢
河南医学研究(2022年19期)2022-10-19
中国比较医学杂志(2020年4期)2020-05-26
水生生物学报(2019年4期)2019-07-20
生态学报(2019年11期)2019-07-08
国际口腔医学杂志(2019年3期)2019-05-31
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
天然产物研究与开发(2018年2期)2018-04-04
医学研究杂志(2015年11期)2015-06-10
食品工业科技(2014年23期)2014-03-11
