
黔中喀斯特次生林主要物种的生态位及种间联结性动态变化
2021-02-05 03:33:06李婷婷容丽王梦洁叶天木王琪
热带亚热带植物学报 2021年1期
李婷婷, 容丽*, 王梦洁, 叶天木, 王琪
黔中喀斯特次生林主要物种的生态位及种间联结性动态变化
李婷婷a, 容丽a*, 王梦洁a, 叶天木a, 王琪b
(贵州师范大学, a. 地理与环境科学学院;b. 喀斯特研究院,贵阳 550025)
为探讨黔中喀斯特次生林演替规律,基于2013-2019年的定点监测数据,从群落物种组成、生态位宽度、生态位重叠值、种间联结等方面研究了黔中喀斯特次生林生态位和种间联结性的动态变化。结果表明,7年间建群种的重要值占比减小,六月雪()、珍珠荚蒾()等物种优势度显著增加。竹叶椒()、小果蔷薇()等占据较宽生态位,云南旌节花()、六月雪等物种对资源的可利用量增加,使得其生态位变窄。生态位重叠值小于0.2的种对比例减少,种间竞争关系更为复杂。主要物种间的总体联结性由不显著正联结变为显著正联结,多数种对间的正负联结比例上升。黔中喀斯特次生林群落中种间竞争激烈且渐趋稳定,种间联结紧密性增强,在正向演替过程中, 伴随着六月雪、珍珠荚蒾等耐阴种和中生性种的优势度和竞争力增大, 小冻绿树()和杭子梢()等先锋种竞争不足而衰退。这为喀斯特地区植被恢复和石漠化治理提供参考。
黔中;喀斯特;群落;生态位;重要值;种间联结性
生态位和种间联结性是种群生态学和群落生态学的重要内容[1],可基于物种对资源的利用能力来解释种间关系特征[2–3],植物群落物种尤其优势物种的生态位和种间关系研究对于理解种间相互关系[4]、群落结构[5]、功能与演替[6]等具有重要作用。生态位的研究主要集中在生态位宽度和生态位重叠的估算和分析上[7–8],种间联结性研究主要是基于二元数据定性或定量分析植物种间的相关性[9],广泛用于各种植被类型的优势种群的种间联结性分析[10]。目前对不同植被类型和种群的生态位和种间联结性动态已有不少研究报道,涉及常绿阔叶林、热带季雨林、针阔混交林、湿地植被、荒漠植被和古树种群等[11–19],但多数仅针对一次性监测数据进行研究,而基于永久样地对自然植被的生态位和种间联结性的研究报道较少。
我国喀斯特地貌集中分布于滇黔桂地区,而贵州地处西南喀斯特生态环境脆弱中心地带,受喀斯特山地植被的特殊生境条件和人为干扰双重影响,区域内石漠化严重,而植被恢复是治理石漠化的有效手段和措施。目前关于喀斯特植被的生态位和种间联结性的研究,侧重于主要物种或优势物种的生态位和种间联结研究[20–22],或以“空间代替时间”的方法[23]进行比较研究, 静态研究较多,对于演替过程中生态位和种间联结性研究较少,尤其是在典型石漠化环境下的植物群落次生演替,其生态位和种间联结性动态研究则鲜有报道,黔中普定县是我国典型的南方喀斯特石漠化严重区域,地形山高坡陡,样地选择困难,由于前期人为干扰,植被受到不同程度的破坏,生态环境严重退化,生境环境脆弱性高、敏感度高,石漠化土地面积占喀斯特出露面积比例大。基于此,以典型的黔中普定县为案例区,基于黔中喀斯特山地次生林群落2013年初测、2015和2019年复测的3次调查数据,研究主要物种的生态位特征和种间关系变化,以植被生态位和种间联结性动态变化来探究黔中喀斯特次生林演替规律,从而为喀斯特地区植被恢复和石漠化治理提供参考。
1 材料和方法
1.1 研究区概况
本研究在普定县(26°22ʹ07.06ʹʹ N, 105°45ʹ06.65ʹʹ E)生态试验站进行,试验站位于贵州省中部偏西,隶属于安顺市,占地总面积11.3 hm2。普定县是典型的喀斯特地貌区,平均海拔为1 176 m,地势为南、北部高,中间低。属于中亚热带季风温暖湿润气候,季风交替十分明显,全年气候温和,辐射能量低,年平均气温15.2℃,年平均日照时数1 189 h,无霜期可达301 d,年平均降水量1 341 mm,降水主要集中于5-9月,相对湿度可达80%。土壤类型以石灰土、黄壤为主。由于人为干扰的影响,原生性的典型喀斯特常绿落叶阔叶混交林已经不存在,群落中藤刺灌丛和灌草丛占优势。调查群落中乔木层以蔷薇科(Rosaceae)、樟科(Lauraceae)、豆科(Legu- minosae)、鼠李科(Rhamnaceae)植物为主,灌木层以蔷薇科、樟科、豆科、桑科(Moraceae)、葡萄科(Vitaceae)、忍冬科(Caprifoliaceae)植物为主, 草本层以禾本科(Poaceae)、莎草科(Cyperaceae)、菊科(Asteraceae)植物为主。
1.2 样地设置和植被调查
为了对喀斯特次生林植被演替进行长期动态监测,以典型南方喀斯特石漠化区普定县的喀斯特山地次生林群落为研究对象,2013年7-8月在普定县城北5 km处的城关镇陇嘎村沙湾组建立了固定样地进行定点监测,样地为典型的喀斯特次生林群落,其基准点地理坐标为26°15′25.8″ N,105°46′ 39.06″ E,海拔为1166~1470 m,以朱守谦等[24]对茂兰喀斯特森林进行大量样地调查所确定的最小表现面积900 m2作为依据,在陈旗地下河小流域内喀斯特坡地上相距200 m的不同方向上分别设置6个不连续样地,每个样地为30 m×30 m,每个样地再细分为36个5 m×5 m的样方。对样方内胸径(DBH)≥1 cm的乔木个体进行挂牌和编号,记录其树高、胸径、冠幅、种名等。对样方内的灌木和草本个体采取系统抽样法,记录其种名、平均高度、株数、平均基径(灌木)、平均盖度(草本)等,并且对木本植物进行每木定位(确定,坐标),同时用GPS记录各样方中心的经纬度和海拔等地理坐标,调查测量其坡度、坡位、坡向、岩石裸露率,根据前期不同人为干扰方式来确定干扰程度,根据各群落整体树高来衡量群落高度。2015年8月和2019年10月采用同样的方法进行复测。
1.3 数据处理
重要值的计算 为了客观反映植物在群落中地位的动态变化情况,分别计算2013、2015和2019年物种的重要值,重要值=(相对密度+相对显著度+相对频度)/3。按照重要值顺序确定对15种主要物种进行生态位和种间联结性动态变化分析。
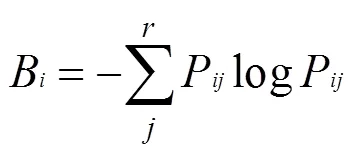
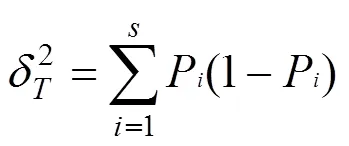
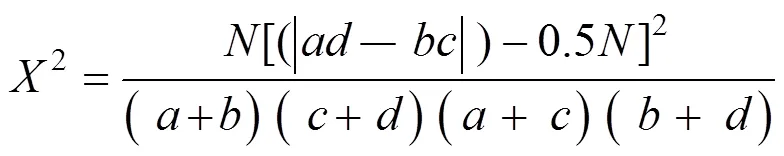
联结系数(AC) 种间关联程度采用联结系数()表示。当≥时,=(-)/(+)(+);当>,
2 结果和分析
2.1 主要物种的重要值变化
2013、2015和2019年调查表明,群落中分别有103种(46科77属)、112种(46科81属) 和102种(48科73属)植物,物种数和个体数量变化显著。从表1可见,3次调查中主要物种的重要值发生变化,2013-2015年建群种由竹叶椒+小冻绿树被圆果化香()+小果蔷薇替代,其中圆果化香的重要值增幅明显(75.55%),但2019年圆果化香又被刺楸()所代替,且重要值减小。重要值排名前2名的比例相差较小, 第1次调查中竹叶椒和小冻绿树重要值占比分别为5.66%和5.24%,第2次调查中圆果化香和小果蔷薇重要值占比分别为8.79%和6%,第3次调查中刺楸和小果蔷薇重要值占比分别为5.49%和5.25%,但重要值比例皆在10%以下。2013-2019年香椿()、珍珠荚蒾、刺楸、朴树()、六月雪的重要值增加,其中刺楸增加了约3.5倍。珍珠荚蒾和六月雪是中生性偏阴和耐阴物种,其重要值和排序在演替过程中均有明显增加。竹叶椒、火棘()、杭子梢、槲栎()、红毛悬钩子()、灰毛浆果楝()的重要值在3次调查中均有不同幅度的下降,优势地位在减弱。
2.2 主要物种的生态位宽度变化
由表1可见,3次调查中主要物种的生态位宽度均有变化,珍珠荚蒾和小果蔷薇的生态位宽度值均大于0.6,在群落中可利用的资源较丰富,生态幅较广,对环境的利用能力较强。2013-2019年竹叶椒、小冻绿树、小果蔷薇、圆果化香、槲栎、火棘、香椿和杭子梢的生态位宽度值显著增加,在群落中的资源利用及其对环境的适应能力逐渐增强,其中圆果化香虽优势地位明显,但占据较窄的生态位宽度,3次调查的生态位宽度值皆小于0.5。灰毛浆果楝、盐肤木()的生态位宽度值皆小于0.4,占据较窄的生态位宽度,对资源环境的利用能力较弱。云南旌节花、六月雪、盐肤木、刺楸、朴树和珍珠荚蒾等6物种生态位宽度值7年间减少,其生态适应范围在不断减小,对资源环境的利用能力逐渐减弱,其中朴树的生态位宽度降幅最大(43.03%),珍珠荚蒾降幅最小(4.11%)。
2.3 主要物种的生态重叠值动态变化
群落中3次调查的主要物种形成105个种对,其中生态重叠值大于0.5和小于0.5的比例基本维持稳定。7年间群落中生态位重叠值小于0.2的物种种对明显减少,从17对下降为10对,占比从9.94%减少到5.84%,且群落中未出现生态位无重叠的种对,表明在黔中喀斯特次生林中主要物种间普遍存在生态位重叠现象,主要物种能对周边资源环境进行充分利用。小果蔷薇和小冻绿树的生态位重叠值在7年间均较高,表明这2种物种在群落中对资源环境的竞争较大,物种生态位趋于泛化。
由表2可看出,7年间主要物种种对的生态位重叠值大部分呈现增加趋势,其中重叠值增加的种对有91对,减少的有80对,平均值从0.524增加到0.558,物种竞争随时间不断增强。3次调查发现阳生性物种间的生态位重叠值比耐阴物种与阳生性物种的重叠值大,如小果蔷薇-杭子梢比小果蔷薇-六月雪的重叠值高。阳生性物种大部分的重叠值仍在增加,如小果蔷薇、小冻绿树与其他种群的种对,中生性物种与其他种群的重叠值大部分略微增加,如槲栎、珍珠荚蒾和朴树,阴生性物种与其他种群的重叠值大部分也呈增加趋势,如地果、六月雪和朴树。群落中小果蔷薇的生态位较宽,与竹叶椒、杭子梢等物种的生态位重叠值大于0.6;槲栎的生态位宽度较窄,与香椿、圆果化香、云南旌节花等生态位窄的物种生态位重叠值也较小。同时也存在生态位较窄物种小冻绿树与生态位较宽物种圆果化香的生态位重叠值均大于0.6,这表明生态位重叠与生态位宽度是密切相关的。圆果化香除了与香椿、火棘和朴树的生态位重叠值减少,其他种对皆逐渐增加。小冻绿树与其他物种的生态位重叠值多数虽大于0.6,但7年间与竹叶椒、圆果化香、六月雪、盐肤木等物种的生态位重叠值呈现逐渐减少趋势,杭子梢除了与圆果化香、香椿和槲栎的生态位重叠值增加,与其他物种的生态位重叠值减少。
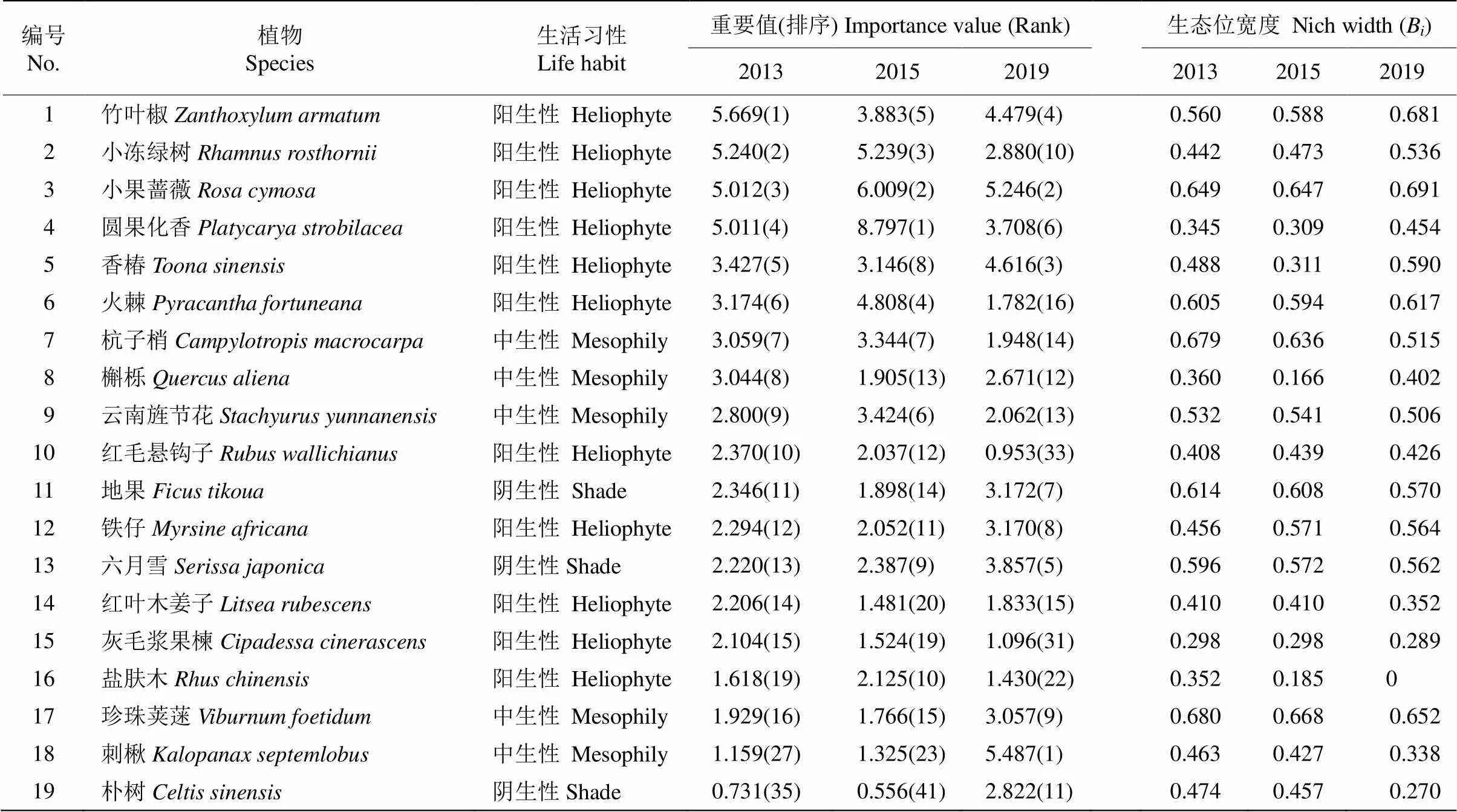
表1 2013-2019年主要物种的重要值和生态位宽度变化
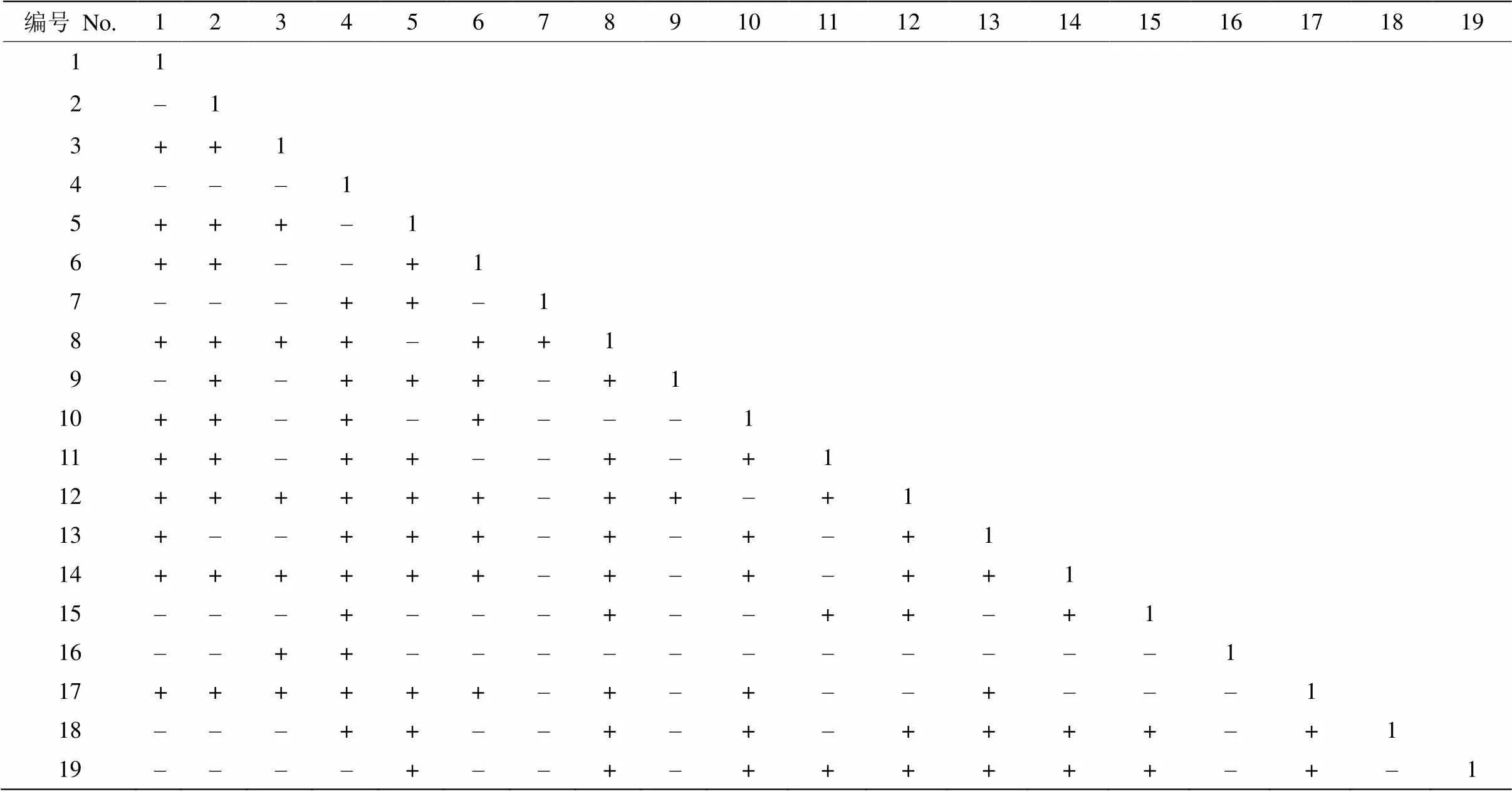
表2 2013-2019年黔中喀斯特次生林主要物种间的重叠值变化
编号见表1; +: 增加; -: 减少。
No. see Table 1+: Increase; -: Decrease.
2.4 主要物种的种间联结动态
黔中喀斯特次生林群落主要物种的总体关联性见表3,3次调查的主要物种间总体关联性由不显著正关联变为显著正关联,黔中喀斯特次生林群落物种的方差比率()分别为1.3、1.77和2.47,呈现增加趋势且皆大于1,表现为正联结, 检验统计量分别为6.5、8.83和12.36,2013-2015年总体联结性皆表现为不显著正联结,然而2019年的>20.05(N),总体联结性表现为显著正联结, 这表明群落中主要物种间的总体联结性增强。
种间联结的2检验结果表明(图1),主要物种组成的105个种对中,2013-2015年种间联结变化不显著,基本稳定,正联结从20对减少到17对,占总种对数比从19.04%减少到16.19%,其中显著正联结从1对增加到2对,但2019年的调查表明,正联结增加到26对,占总种对数的24.76%,其中显著正联结6对。7年间负联结种对数先减少后增加,从15对减至11对后又增至19对,分别占总种对数的14.28%、10.47%和18.09%。第1次调查中只有竹叶椒和云南旌节花呈显著正关联(3.841<2<6.635)。小冻绿树和六月雪以及云南旌节花和铁仔()这2对种对在第2次调查中呈显著正关联,第3次调查中小冻绿树与云南旌节花、小冻绿树与铁仔、刺楸与圆果化香、杭子梢与六月雪、朴树与槲栎以及云南旌节花与铁仔的种间联结皆呈显著正联结。但所有种对中多数种对呈现不显著正或负关联和无联结性,说明在群落中的主要物种间的种间联结性较松散,种对间对生境和生物学特性的差异较大、种对间独立性较强。

表3 2013-2019年主要物种间的总体关联性
种间联结的联结系数(AC)结果表明(图2),7年间105对种对中正联结的种对数呈现先减少后增加趋势,由20对减至16对又增至25对,负联结的种对数呈现先减少后增加趋势,由15对减至10对又增至19对,无联结(AC=0)的种对数呈增加趋势,从25对增至55对。由于有一些物种出现在所有样地中,所以无法计算其AC值的种对逐渐减少。-0.6≤AC≤0.6的种对从10对减少到6对,占比从9.52%减少到5.71%,说明这些种对间的正联结明显,具有非常相似的生态适应性。–1≤AC≤-0.6的种对从12对减少到10对又增至16对,说明这些种对的负联结明显,生态适应性差异明显,存在着竞争。所有的显著正负联结的AC值都为1或-1,这与两物种没有共同出现的样地相关,因此还需结合2的检验来进行分析。
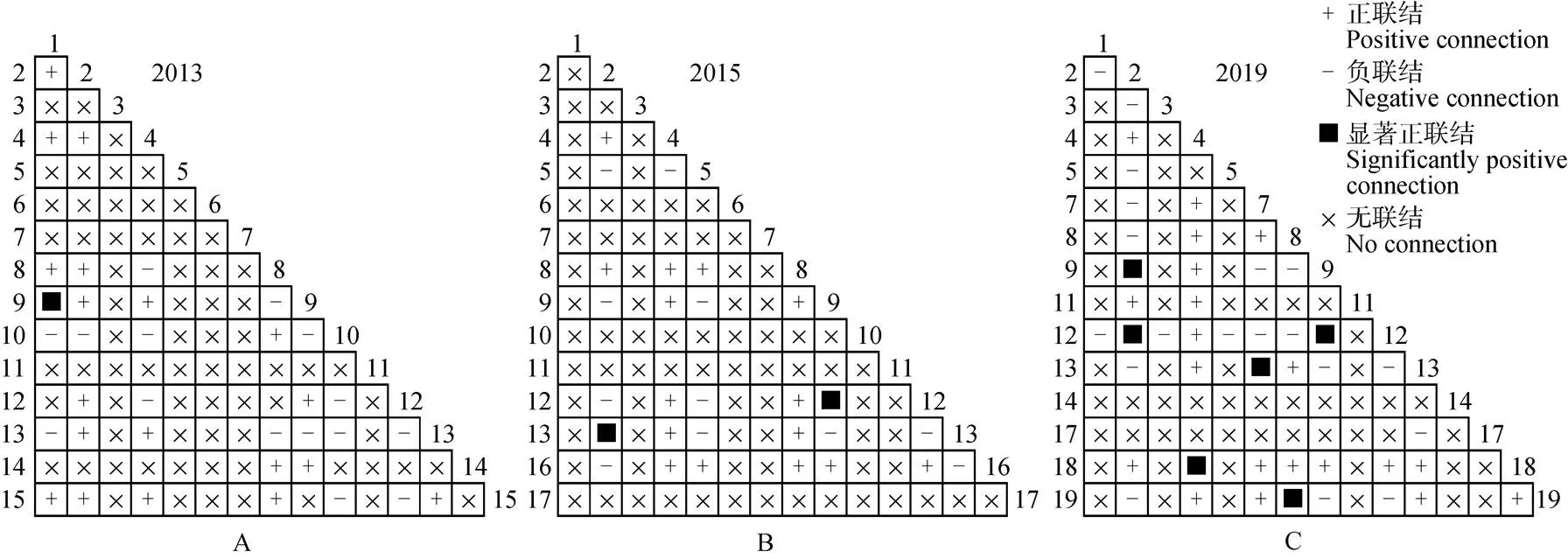
图1 2013-2019年主要物种间X2检验半矩阵图。1~19见表1。
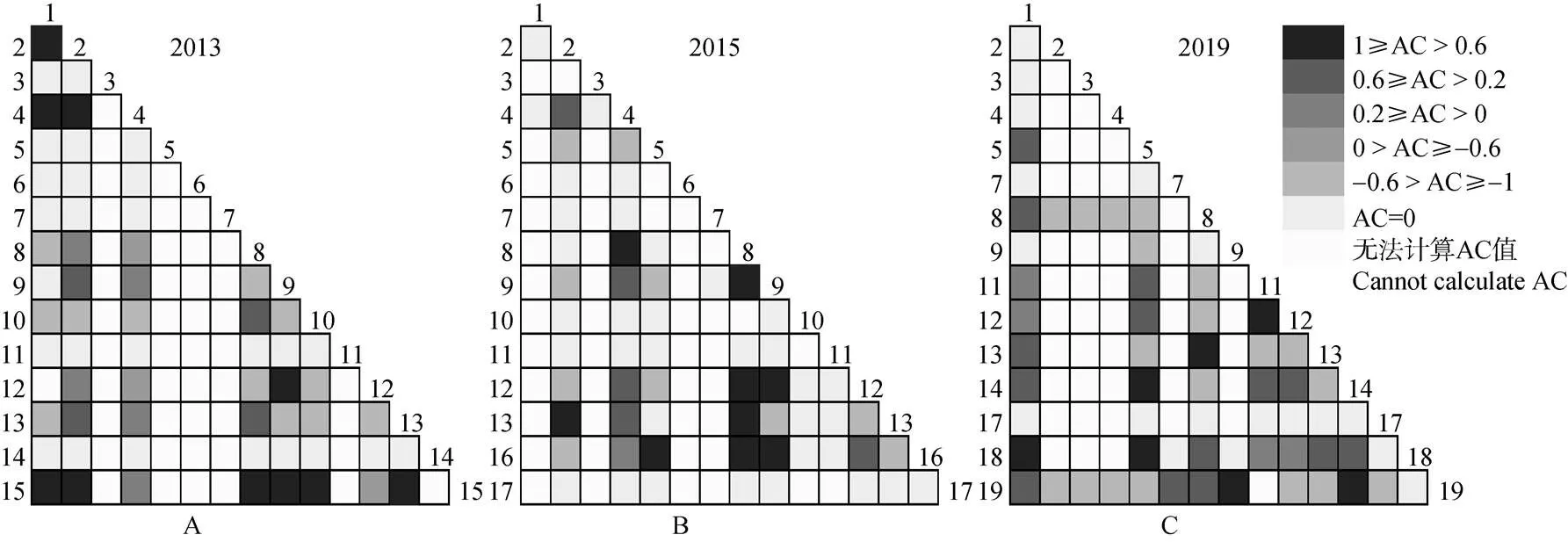
图2 2013-2019年主要物种间种间联结半矩阵图。1~19见表1。
3 结论和讨论
3.1 建群种重要值不高,耐荫和中生性物种的重要值逐渐增大
2013、2015和2019年的调查结果表明,黔中喀斯特次生林是由多个种群组成的复杂群落,群落中没有非常明显的优势种,群落中建群种竹叶椒+小冻绿树被圆果化香+小果蔷薇替代,但在2019年又被小果蔷薇+刺楸代替,物种的重要值减小且占比小,优势度不明显。喻理飞等[27]在对茂兰自然保护区的树种适应等级种组划分研究中,将圆果化香定义为顶极种;李援越等[28]对黔中喀斯特次生林研究中将其定义为次顶极种。本研究中圆果化香2015年的重要值增幅最大(4.07%),但是在2019年重要值下降,表明演替顶极种圆果化香在群落中已开始占主要地位,圆果化香是阳生性树种,具有光补偿点、饱和点高,耐旱性强的特点[29],其地上部分芽、枝、叶等构件拓展其生存空间,提高光合面积,地下部分通过根系在岩石缝隙中穿透力强,提高吸收面积,从而提高对恶劣环境的生存适应与竞争能力[29],说明圆果化香的生长能够适应特殊的喀斯特生境,演替过程中土壤养分增加,光热条件不断改善,其种群高度、盖度、显著度不断增加,物种结构渐趋复杂,在演替过程中得到稳定发展,但由于研究区域靠近人口聚集地,可能受到人为干扰使圆果化香的重要值呈现下降趋势。小果蔷薇、竹叶椒等阳生性先锋种的重要值在群落中排名靠前,说明阳生性先锋物种数量多,在群落中仍然处于良好生长阶段。群落中珍珠荚蒾、刺楸和六月雪等中生性偏耐阴和耐阴种的重要值略微增加,林分郁闭度增大,林下层所接收的光照较少,为耐阴性植物提供较好的生境条件,黔中喀斯特植被群落在演替的进程中逐渐出现更多耐阴种,预示着群落可能处于演替中前期,并且正在往正向演替方向发展,这与杨华斌[30]对黔中喀斯特演替阶段的研究结果有相似之处。
3.2 建群种和伴生种的生态位宽度变化
生态位宽度表示物种在生长过程中综合利用资源的能力、竞争水平和对环境的适应能力[31]。7年间竹叶椒、小冻绿树、小果蔷薇和香椿的生态位宽度明显增加,物种数量多,分布广泛,对资源环境利用能力较强,能够适应土层浅薄的喀斯特次生林特殊生境。3次调查中小果蔷薇宽度值均较高,说明喀斯特早中期先锋种较早在群落中占据空间并定居,演替中前期竞争力仍强,但在演替过程中会被喀斯特次顶极和顶极种淘汰,群落会逐渐从结构简单的群落向结构功能复杂的稳定群落发展,与司彬[32]对黔中喀斯特植被自然演替的研究结果部分相似。有研究表明演替早期阶段中不但存在先锋种,而且也存在演替系列群落种和顶极群落种,往往在演替过程中,个体较小、生长较快、寿命较短的种逐渐被个体较大、生长较慢、寿命较长的种取代[33],3次调查中,喀斯特次顶极种圆果化香的重要值高而生态位宽度相对较窄,但会随着演替进展逐渐增加,这可能是喀斯特次生林群落向正向演替发展的重要特征,圆果化香正处于良好发展趋势,在样地中的分布比较集中。对小五台山温带植被[34]、喀斯特季节性雨林[35]、武夷山常阔叶林[36]和茂兰喀斯特森林[23]的优势种群生态位研究也有相似结果。云南旌节花、六月雪、盐肤木、刺楸和朴树的生态位宽度逐渐变窄,在演替过程中, 群落中土壤厚度不断积累,群落郁闭度增大,土壤湿度增加等环境条件的改善,从而对资源利用的选择性逐渐增强,并且对生态因子需求的专化程度增加,其资源的可利用量会逐渐增加[37]。
3.3 种间竞争复杂且激烈,演替中前期种将替代群落先锋种
生态位重叠是指2种或2种以上生态位相似的物种生活于同一空间时,分享或竞争共同资源的现象[38]。本研究中,7年间总体上群落中生态位重叠值小于0.2的种对占比明显在减少,以及生态位重叠值排名前三的种对总体上重叠值高但随演替逐渐减少,群落中小果蔷薇与杭子梢等阳生性种的生态位重叠值大于其与六月雪等耐阴种的生态位重叠值,说明物种在群落中因其高度的生态相似性或者生态互补从而形成资源利用性竞争,在资源环境利用上有所分化,而阳生性物种的重叠值仍在增加,而中生性和阴生性物种呈现略微增加趋势,说明群落阳性先锋种在群落中种间竞争仍激烈,而中生性和阴生性物种随演替竞争表现增强趋势。多数研究认为,较大生态位宽度往往伴随着较高的生态位重叠值[39–40],如本研究中的小果蔷薇和杭子梢。黔中喀斯特植被的生态位重叠值和生态位宽度之间关联密切,但也存在矛盾的情况,生态位宽度窄的种对生态位重叠值小,如槲栎和圆果化香;生态位较窄与生态位较宽的种对间重叠值高,如小冻绿树与圆果化香,可能是黔中喀斯特次生林在演替过程中环境资源存在着高度的空间异质性,以及群落中组成结构不稳定所造成的,说明群落种间竞争复杂且激烈,但在演替进展中会逐渐趋于稳定,这与覃弦等[23]对茂兰乔木层不同演替阶段的生态位研究结论相似,说明在喀斯特植被演替中具有相似的规律。珍珠荚蒾、六月雪和圆果化香等演替中前期物种在演替过程中个体数量增加,生态位重叠值增大,竞争力增强而逐渐淘汰竞争力不足的喀斯特先锋种,如杭子梢、小冻绿树。
3.4 植被群落趋于稳定,种间联结性虽松散,但紧密性增强
随着植物群落的演替,林龄增长,群落内物种的生态位分化越来越合理,种间关系趋于正联结[41],2013、2015和2019年调查结果表明,主要物种之间的总体种间关系表现为由不显著正联结变为显著正联结,说明群落结构及其组成成分随演替趋于稳定。本研究中呈现显著正联结的种对数在7年间呈现增加趋势,由1对增加至6对,表明种对间的关系逐渐紧密,生态习性相似,具有互补的生境要求,这与桂西南喀斯特山地的种间关系研究有相似结论[42]。2检验和AC值表明7年间多数种对间呈现无关联和不显著正负关联的情况,但是在第三次调查中正负联结比例均趋于增加,说明主要物种间联结性较为松散,多数物种间的种间独立性较强,但一定程度上主要物种间紧密性趋于加强,但主要物种能够占据有利于自身的生态位的同时也可能受到了前期人为干扰的影响,表明群落演替到中前期阶段,仍需加强人为保护措施,在刘润红等[43]和陈玉凯等[44]的种间关系研究中有相似结论,同时也可能与所调查样地面积、生物生态习性和优势种的变化有关。可能预示着群落在演替过程中趋于稳定,喀斯特次生林在恢复过程中往正向演替方向发展。
本研究结果表明,7年间建群种重要值皆不高,而珍珠荚蒾、六月雪、刺楸的重要值逐渐增加;竹叶椒、小果蔷薇、小冻绿树和香椿占据较宽的生态位,资源的利用能力强,而云南旌节花、六月雪、盐肤木、刺楸和朴树的资源可利用量逐渐增加,使其生态位宽度变窄;生态位重叠值小于0.2的种对占比明显减少,种间竞争关系渐趋激烈复杂,群落中生态位宽度与重叠值关联显著,圆果化香、珍珠荚蒾和六月雪等物种的生态位重叠增大,在演替过程中逐渐淘汰重叠值减小的先锋种,如杭子梢和小冻绿树;群落中总体关联性由不显著正关联变化为显著正关联,无关联和不显著正负关联的种对较多,正负关联种对比例增加,说明植被群落会逐渐趋于稳定,种间联结虽松散,但其紧密性趋于增强。
黔中次生林群落演替中,群落逐渐稳定,种间联结松散,在正向演替过程中伴随着六月雪和珍珠荚蒾等耐荫种和中性种优势地位和竞争力的增加以及杭子梢和小冻绿树等先锋种的淘汰。黔中地区植被的种间竞争在自然演替过程中是稳定朝着正向演替的方向发展的,应采取相应的措施减少人为干扰,加强保护力度来防止植被再次退化。
[1] GRINNELL J. The niche-relationships of the California thrasher [J]. Auk, 1917, 34(4): 427–433. doi: 10.2307/4072271.
[2] YANG Z H, LANG X D, LI S F, et al. Niche and interspecific inter- action of dominant tree population incommunities[J]. For Res, 2015, 28(4): 473–478. doi: 10.3969/j.issn.1001-1498.2015. 04.004. 杨宗慧, 郎学东, 李帅锋, 等. 篦子三尖杉群落优势种群生态位和种间关系[J]. 林业科学研究, 2015, 28(4): 473–478. doi: 10.3969/j. issn.1001-1498.2015.04.004.
[3] DENG X L, CAO Y S, LIANG Q, et al. Study on niche of dominant species at arbor layer in montane elfin forest of Jinggangshan [J]. J Plant Res Environ, 2016, 25(1): 88–93. doi: 10.3969/j.issn.1674-7895. 2016.01.11. 邓贤兰, 曹裕松, 梁琴, 等. 井冈山山顶矮林乔木层优势种的生态位研究[J]. 植物资源与环境学报, 2016, 25(1): 88–93. doi: 10.3969/ j.issn.1674-7895.2016.01.11.
[4] HAN J T, LI S Q, ZHAO D H, et al. Interspecific relationships among dominant species in the planted vegetation communities in sandy hill regions of northwestern Shanxi, China [J]. J Arid Land Res. Environ, 2016, 30(12): 164–169. doi: 10.13448/j.cnki.jalre.2016.400. 韩锦涛, 李素清, 赵德怀, 等. 晋西北丘陵风沙区人工植物群落优势种种间关系研究[J]. 干旱区资源与环境, 2016, 30(12): 164–169. doi: 10.13448/j.cnki.jalre.2016.400.
[5] ZHANG M X, WANG D X, KANG B, et al. Interspecific associations of dominant plant populations in secondary forest ofin Qinling Mountains [J]. Sci Silv Sin 2015, 51(1): 12–21. doi: 10.11707/ j.1001-7488.20150102. 张明霞, 王得祥, 康冰, 等. 秦岭华山松天然次生林优势种群的种间联结性[J]. 林业科学, 2015, 51(1): 12–21. doi: 10.11707/j.1001- 7488.20150102.
[6] KANG Y X, KANG B W, YUE J W, et al. The classification ofcommunities and their niche in Loess Plateau of north Shaanxi [J]. Acta Ecol Sin, 2007, 27(10): 4096–4105. doi: 10. 3321/j.issn:1000-0933.2007.10.017. 康永祥, 康博文, 岳军伟, 等. 陕北黄土高原辽东栎()群落类型划分及其生态位特征[J]. 生态学报, 2007, 27 (10): 4096–4105. doi: 10.3321/j.issn:1000-0933.2007.10.017.
[7] LI S F, LIU W D, SU J R, et al. Niches and interspecific associations of dominant tree populations at different restoration stages of monsoonal broad-leaved evergreen forest [J]. Chin J Ecol, 2011, 30(3): 508–515. doi: 10.13292/j.1000-4890.2011.0072. 李帅锋, 刘万德, 苏建荣, 等. 季风常绿阔叶林不同恢复阶段乔木优势种群生态位和种间联结[J]. 生态学杂志, 2011, 30(3): 508–515. doi: 10.13292/j.1000-4890.2011.0072.
[8] LU W L, ZHANG H J, CHEN Q B, et al. Niche characteristics of the tree-layer dominant populations offorest in Chongqing Simian Mountains [J]. J NW For Univ, 2016, 31(2): 60–65. doi: 10.3969/j.issn.1001-7461.2016.02.11. 卢炜丽, 张洪江, 陈奇伯, 等. 重庆四面山杉木林群落乔木层优势种群生态位特征研究[J]. 西北林学院学报, 2016, 31(2): 60–65. doi: 10.3969/j.issn.1001-7461.2016.02.11.
[9] XU M H, LIU M, ZHAI D T, et al. A review of contents and methods used to analyze various aspects of plant interspecific associations [J]. Acta Ecol Sin, 2016, 36(24): 8224–8233. doi: 10.5846/stxb2015053 11092. 徐满厚, 刘敏, 翟大彤, 等. 植物种间联结研究内容与方法评述[J]. 生态学报, 2016, 36(24): 8224–8233. doi: 10.5846/stxb201505311092.
[10] PAN G, ZHANG H P, PAN D. Interspecific associations of dominant plant populations in a mid-subtropicalforest [J]. Chin J Ecol, 2017, 36(4): 892–901. doi: 10.13292/j.1000-4890. 201704.020.潘高, 张合平, 潘登. 中亚热带南酸枣林优势种群的种间联结性[J]. 生态学杂志, 2017, 36(4): 892–901. doi: 10.13292/j.1000-4890. 201704.020.
[11] YE D, WEN Y G, DENG R Y, et al. Dynamic changes of plant population niche in succession series of evergreen broad-leaved forest in Daming Mountain of Guanxi [J]. Chin J Ecol, 2009, 28(3): 417–423. 叶铎, 温远光, 邓荣艳, 等. 大明山常绿阔叶林演替序列种群生态位动态特征[J]. 生态学杂志, 2009, 28(3): 417–423.
[12] ZHANG G M, TANG J W, SHI J P, et al. Niche dynamics of dominant populations ofColia pioneer community in Xishuang- banna, SW. China [J]. J Plant Res Environ, 2000, 9(1): 22–26. doi: 10. 3969/j.issn.1674-7895.2000.01.006. 张光明, 唐建维, 施济普, 等. 西双版纳野芭蕉先锋群落优势种群的生态位动态[J]. 植物资源与环境学报, 2000, 9(1): 22–26. doi: 10. 3969/j.issn.1674-7895.2000.01.006.
[13] YANG Q, ZHOU J, TAO C, et al. Comparative study on interspecific association in tropical evergreen monsoon elfin forest at two succe- ssional stages in Tongguling of Hainan Island [J]. Chin Agric Sci Bull, 2014, 30(22): 8–15. 杨琦, 周婧, 陶楚, 等. 海南岛铜鼓岭热带常绿季雨矮林2个演替阶段的种间联结性对比研究[J]. 中国农学通报, 2014, 30(22): 8–15.
[14] ZOU S, ZHOU G Y, ZHANG Q M, et al. Long-term (1992-2015) dynamics of interspecific associations among tree species in a monsoon evergreen broad-leaved forest in Dinghushan Biosphere Reserve [J]. Acta Ecol Sin, 2019, 39(17): 6362–6371. doi: 10.5846/stxb20180403 0753. 邹顺, 周国逸, 张倩媚, 等. 1992-2015年鼎湖山季风常绿阔叶林群落种间关联动态[J]. 生态学报, 2019, 39(17): 6362–6371. doi: 10. 5846/stxb201804030753.
[15] FU W G, LI P P, BIAN X M, et al. Niche dynamics of populations of vegegation community in succession process in Zhenjiang Inner-river wetland [J]. Ecol Environ, 2008, 17(1): 278–284. doi: 10.16258/j.cnki. 1674-5906.2008.01.047. 付为国, 李萍萍, 卞新民, 等. 镇江内江湿地植物群落演替进程中种群生态位动态 [J]. 生态环境, 2008, 17(1): 278–284. doi: 10. 16258/j.cnki.1674-5906.2008.01.047.
[16] GONG Z W, KANG X G, GU L, et al. Interspecific association among arbor species in two succession stages of spruce-fir conifer and broad- leaved mixed forest in Changbai Mountains, northeastern China [J]. J Beijing For Univ, 2011, 33(5): 28–33. doi: 10.13332/j.1000-1522.2011. 05.018. 龚直文, 亢新刚, 顾丽, 等. 长白山云冷杉针阔混交林两个演替阶段乔木的种间联结性 [J]. 北京林业大学学报, 2011, 33(5): 28–33. doi: 10.13332/j.1000-1522.2011.05.018.
[17] LAAKA-LINDBERG S, POHJAMO M, KORPELAINEN H. Niche breadth and niche overlap in three epixylic hepatics in a boreal old- growth forest, southern Finland [J]. J Bryol, 2005, 27(2): 119–127. doi: 10.1179/037366805X53031.
[18] MYSTER R W, PICKETT T A. Dynamics of associations between plants in ten old fields during 31 years of succession [J]. J Ecol, 1992, 80(2): 291–302. doi: 10.2307/2261012.
[19] CHEN Y R, YIN L K. Community composition and niche change characteristics of dominant species in the wind-breaking and sand- fixing forest, Xinjiang, China [J]. J Plant Ecol, 2008, 32(5): 1126–1133. doi: 10.3773/j.issn.1005-264x.2008.05.017. 陈艳瑞, 尹林克. 人工防风固沙林演替中群落组成和优势种群生态位变化特征 [J]. 植物生态学报, 2008, 32(5): 1126–1133. doi: 10. 3773/j.issn.1005-264x.2008.05.017.
[20] QIN S T, LONG C L, WU B L. Study on niche of dominant shrub populations in karst forest in Maolan Nature Reserve, Guizhou Province [J]. For Resour Manage, 2018(2): 58–64,90. doi: 10.13466/j. cnki.lyzygl.2018.02.011. 秦随涛, 龙翠玲, 吴邦利. 茂兰喀斯特森林优势灌木种群的生态位研究[J]. 林业资源管理, 2018(2): 58–64,90. doi: 10.13466/j.cnki. lyzygl.2018.02.011.
[21] YU X Y, YU R, HUANG J, et al. Analysis on interspecific associations among dominant populations in each layer ofcommunity in Maolan National Nature Reserve of Guizhou [J]. J Plant Res Environ, 2018, 27(2): 24–32. doi: 10.3969/j.issn.1674- 7895.2018.02.04. 俞筱押, 余瑞, 黄娟, 等. 贵州茂兰国家级自然保护区四药门花群落各层次优势种群的种间联结性分析[J]. 植物资源与环境学报, 2018, 27(2): 24–32. doi: 10.3969/j.issn.1674-7895.2018.02.04.
[22] ZHANG Z H, HU G. Interspecific relationships of dominant species incommunity in karst mountain area [J]. Ecol Environ Sci, 2011, 20(8/9): 1209–1213. doi: 10.3969/j.issn.1674-5906. 2011.08.004. 张忠华, 胡刚. 喀斯特山地青冈栎群落优势种的种间关系分析[J]. 生态环境学报, 2011, 20(8/9): 1209–1213. doi: 10.3969/j.issn.1674- 5906.2011.08.004.
[23] QIN X, LONG C L, LI J. Analysis of ecological niches of dominant arbor populations in different succession stages in the karst forest in Maolan Nature Reserve [J]. J SW Univ (Nat Sci), 2017, 39(6): 42–49. doi: 10.13718/j.cnki.xdzk.2017.06.007. 覃弦, 龙翠玲, 李娟. 茂兰喀斯特森林不同演替阶段优势乔木种群生态位研究[J]. 西南大学学报(自然科学版), 2017, 39(6): 42–49. doi: 10.13718/j.cnki.xdzk.2017.06.007.
[24] ZHU S Q, CHEN Z R, WEI L M. The process and pattern of natural restoration in degraded Karst forest [J]. J Guizhou Univ (Agric Biol Sci), 2002, 21(1): 19–25. doi: 10.3969/j.issn.1008-0457.2002.01.005. 朱守谦, 陈正仁, 魏鲁明. 退化喀斯特森林自然恢复的过程和格局[J]. 贵州大学学报(农业与生物科学版), 2002, 21(1): 19–25. doi: 10. 3969/j.issn.1008-0457.2002.01.005.
[25] LEVINS R. Evolution in Changing Environments: Some Theoretical Explorations [M]. Princetion: Princetion University Press, 1968.
[26] SCHLUTER D. A variance test for detecting species associations with some example applications [J]. Ecology, 1984, 65(3): 998–1005.
[27] YU L F, ZHU S Q, YE J Z, et al. Dynamics of a degraded Karst forest in the process of natural restoration [J]. Sci Silv Sin, 2002, 38(1): 1–7. doi: 10.3321/j.issn:1001-7488.2002.01.001. 喻理飞, 朱守谦, 叶镜中, 等. 退化喀斯特森林自然恢复过程中群落动态研究[J]. 林业科学, 2002, 38(1): 1–7. doi: 10.3321/j.issn: 1001-7488.2002.01.001.
[28] LI Y Y, ZHU S Q, ZHU X K. Number classification of degraded karst forests in the central Guizhou [J]. J Mount Agric Biol, 2000, 19(2): 94– 98. doi: 10.3969/j.issn.1008-0457.2000.02.004. 李援越, 朱守谦, 祝小科. 黔中退化喀斯特森林植物群落的数量分类[J]. 山地农业生物学报, 2000, 19(2): 94–98. doi: 10.3969/j.issn. 1008-0457.2000.02.004.
[29] YU L F. Primary study on the structure of bud population for[J]. Guizhou Sci, 2003, 21(2): 64–68. doi: 10.3969/j. issn.1003-6563.2003.01.015. 喻理飞. 圆果化香芽种群结构初步研究[J]. 贵州科学, 2003, 21(2): 64–68. doi: 10.3969/j.issn.1003-6563.2003.01.015.
[30] YANG H B, WEI X L, DANG W. Species composition and diversity variations at different succession stages of karst vegetation in central Guizhou [J]. J Mount Agric Biol, 2009, 28(3): 203–207. doi: 10.3969/j. issn.1008-0457.2009.03.004. 杨华斌, 韦小丽, 党伟. 黔中喀斯特植被不同演替阶段群落物种组成及多样性[J]. 山地农业生物学报, 2009, 28(3): 203–207.
[31] LIU J Z, CHEN Y N, ZHANG Y M. Niche characteristics of plants on four environmental gradients in middle reaches of Tarim River [J]. Chin J Appl Ecol, 2004, 15(4): 549–555. 刘加珍, 陈亚宁, 张元明, 塔里木河中游植物种群在四种环境梯度上的生态位特征[J]. 应用生态学报, 2004, 15(4): 549–555
[32] SI B, YAO X H, REN H D, et al. Species composition and diversity in the process of natural succession of Karst vegetation in central Guizhou: Case study of Puding country in Guizhou [J]. For Res, 2008, 21(5): 669–674. doi: 10.3321/j.issn:1001-1498.2008.05.014. 司彬, 姚小华, 任华东, 等. 黔中喀斯特植被自然演替过程中物种组成及多样性研究——以贵州省普定县为例[J]. 林业科学研究, 2008, 21(5): 669–674. doi: 10.3321/j.issn:1001-1498.2008.05.014.
[33] SONG Y C. Vegetation Ecology [M]. Shanghai: East China Normal University Press, 2001: 267–268. 宋永昌. 植被生态学[M]. 上海: 华东师范大学出版社, 2001: 267– 268.
[34] BAI X H, ZHANG J T. Niche analysis of dominant species of forest community in Xiaowutai Mountain, China [J]. Chin J Appl Ecol, 2017, 28(12): 3815–3826. doi: 10.13287/j.1001-9332.201712.002. 白晓航, 张金屯. 小五台山森林群落优势种的生态位分析[J]. 应用生态学报, 2017, 28(12): 3815–3826. doi: 10.13287/j.1001-9332. 201712.002.
[35] HUANG F Z, LI D X, WANG B, et al. Niche characteristics of dominant populations of tropical karst seasonal rainforest implications for vegetation restoration in rock desertification region [J]. Guangxi Sci, 2018, 25(5): 599–610. doi: 10.13656/j.cnki.gxkx.20181030.006. 黄甫昭, 李冬兴, 王斌, 等. 喀斯特季节性雨林优势种群生态位特征及其对石漠化地区植被修复的启示[J]. 广西科学, 2018, 25(5): 599–610. doi: 10.13656/j.cnki.gxkx.20181030.006.
[36] CHEN X. Analysis of niches for several dominant tree species in evergreen broadleaved forest in Wuyishan [J]. J For Environ, 2016, 36(3): 337–341. doi: 10.13324/j.cnki.jfcf.2016.03.014. 陈晓. 武夷山常绿阔叶林若干优势树种的生态位分析[J]. 森林与环境学报, 2016, 36(3): 337–341. doi: 10.13324/j.cnki.jfcf.2016.03.014.
[37] QIN L. Statistical Ecology [M]. Beijing: China Forestry Press, 2009: 84–85. 覃林. 统计生态学[M]. 北京: 中国林业出版社, 2009: 84–85.
[38] YANG N, ZOU D S, LI J G, et al. Niche dynamics of main plant communities in natural restoration succession process on sloping land with purple soils in Hengyang Basin [J]. Bull Soil Water Conserv, 2010, 30(4): 87–93. doi: 10.13961/j.cnki.stbctb.2010.04.004. 杨宁, 邹冬生, 李建国, 等. 衡阳盆地紫色土丘陵坡地主要植物群落自然恢复演替进程中种群生态位动态 [J]. 水土保持通报, 2010, 30(4): 87–93. doi: 10.13961/j.cnki.stbctb.2010.04.004.
[39] ZHU D H, LIU J F, HONG W, et al. Studies on Regeneration niche of main species incommunity in Daiyun Mountain [J] J Trop Subtrop Bot, 2012, 20(6): 561–565. doi: 10.3969/j.issn.1005- 3395.2012.06.004. 朱德煌, 刘金福, 洪伟, 等. 戴云山黄山松群落主要树种更新生态位研究 [J]. 热带亚热带植物学报, 2012, 20(6): 561–565. doi: 10. 3969/j.issn.1005-3395.2012.06.004.
[40] DONG D, XU X T, ZHOU Z X, et al. Niche dynamics of main populations of old-tree communities in Jiuhua Mountain Scenic Area of Anhui Province [J]. Chin J Ecol, 2019, 38(5): 1292–1304. doi: 10. 13292/j.1000-4890.201905.023. 董冬, 许小天, 周志翔, 等. 安徽九华山风景区古树群落主要种群生态位的动态变化 [J]. 生态学杂志, 2019, 38(5): 1292–1304. doi: 10.13292/j.1000-4890.201905.023.
[41] PENG S L, YAN X T, QI G, et al. Interspecific association dynamic changes among the dominant species ofcommunity in Funiushan National Nature Reserve [J]. J For Environ, 2018, 38(3): 326–333. doi: 10.13324/j.cnki.jfcf.2018.03.011. 彭舜磊, 闫小婷, 齐光, 等. 伏牛山栓皮栎群落优势种群种间关联动态分析 [J]. 森林与环境学报, 2018, 38(3): 326–333. doi: 10. 13324/j. cnki.jfcf.2018.03.011.
[42] MA F F, PAN G, LI X Q, et al. Interspecific relationship and canonical correspondence analysis within woody plant communities in the karst mountains of Southwest Guangxi, southern China [J]. J Beijing For Univ, 2017, 39(6): 32–44. doi: 10.13332/j.1000-1522.20160379. 马丰丰, 潘高, 李锡泉, 等. 桂西南喀斯特山地木本植物群落种间关系及CCA排序 [J]. 北京林业大学学报, 2017, 39(6): 32–44. doi: 10.13332/j.1000-1522.20160379.
[43] LIU R H, JIANG Y, CHANG B, et al. Interspecific associations and correlations among the main woody plants in acommunity in a riparian zone of Lijiang River, Guilin, Southwest China [J]. Acta Ecol Sin, 2018, 38(19): 6881–6893. doi: 10.5846/stxb 201802270396. 刘润红, 姜勇, 常斌, 等. 漓江河岸带枫杨群落主要木本植物种间联结与相关分析 [J]. 生态学报, 2018, 38(19): 6881–6893. doi: 10. 5846/stxb201802270396.
[44] CHEN Y K, YANG X B, LI D H, et al. Interspecific associations among dominant populations in the communities within Bawangling, Hainan Island [J]. J Trop Subtrop Bot, 2011, 19(3): 237–244. doi: 10.3969/j.issn.1005-3395.2011.03.008. 陈玉凯, 杨小波, 李东海, 等. 海南霸王岭油丹所在群落的优势种群种间联结性研究 [J]. 热带亚热带植物学报, 2011, 19(3): 237–244. doi: 10.3969/j.issn.1005-3395.2011.03.008.
Dynamic Changes in Niche and Interspecific Association of Major Species of Karst Secondary Forest in Central Guizhou
LI Ting-tinga,RONG Lia*,WANG Meng-jiea,YE Tian-mua,WANG Qib
(a. School of Geography and Environmental Sciences; b. School of Karst Science, Guizhou Normal University,Guiyang 550025, China)
To explore the succession rule of karst secondary forest in central Guizhou, based on the fixed-point monitoring data in 2013-2019, the dynamic changes of niche and interspecific association of karst secondary forest in central Guizhou were studied, including community species composition, niche width, niche overlap and interspecific association.The results showed that the proportion of important value of constructive species decreased in 7 years, and the dominance of species, such asand, increased significantly.andoccupied a wide niche, while the niche width ofandand other species have increased resource availability, which narrowed their niche width.The proportion of species pairs with niche overlap less than 0.2 decreased, and the relationship of interspecific competition was more complex.The overall connectivity among main species changed from non-significant positive association to significant positive association, and the proportion of positive and negative association between most species increased.In the karst secondary forest community of central Guizhou, the competition among species was intense and gradually stable, and the connection between species was strengthened.In the process of positive succession, the dominance and competitiveness of shade tolerant and mesophily species, such asandincreased,but some pioneer species, such asand, declined due to insufficient competition. Therefore, these would provide a reference for vegetation restoration and rocky desertification control in karst areas.
Central Guizhou; Karst; Community; Niche; Importance value;Interspecies connectivity
10.11926/jtsb.4252
2020–05–18
2020–06–28
国家十三五重点研发计划课题(2016YFC0502607);贵州省科技计划重大专项([2017]5411);贵州省世界一流学科建设计划项目([2019]125)资助
This work was supported by the Thirtyth Five Program for National Key Research and Development of China (Grant No. 2016YFC0502607), the Key Program for Science and Technology of Guizhou (Grant No. 2017-5411), and the Program for World-class Discipline Construction of Guizhou (Grant No. 2019-125).
李婷婷(1995~ ),女,硕士研究生,主要从事植物地理学研究。E-mail: 1782936282@qq.com
E-mail: ronglit@126.com
猜你喜欢
植物研究(2023年5期)2023-09-09 08:01:22
智慧农业导刊(2022年9期)2022-11-23 13:06:59
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
作文与考试·小学高年级版(2021年5期)2021-03-24 11:37:51
现代养生·下半月(2018年11期)2018-03-20 02:03:22
现代农业研究(2017年11期)2018-01-12 21:03:03
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
现代园艺(2017年20期)2017-02-03 18:49:29
杂草学报(2015年2期)2016-01-04 14:57:55
