
转录调控因子GntR10在布鲁氏菌感染细胞和小鼠模型中的毒力作用研究
2021-01-20 01:21李志强王书利席丽崔艳艳张春梅施传信张辉
商丘师范学院学报 2021年3期
李志强,王书利,席丽,崔艳艳,张春梅,施传信,张辉
(1.商丘师范学院 生物与食品学院,河南 商丘 476000;2.石河子大学 动物科技学院,新疆 石河子 832000)
布鲁氏菌(Brucella)是革兰氏阴性胞内寄生菌,可以在人和动物的专业和非专业吞噬细胞内生存,引起人和多种动物发生疾病[1].在布鲁氏菌的菌属中,羊种布鲁氏菌、猪种布鲁氏菌和牛种布鲁氏菌可以感染人,引起人类布鲁氏菌病[2].牛种布鲁氏菌是牛布鲁氏菌病的病原体,分布广泛,可引起牛的流产和人慢性感染性疾病,特别是在发展中国家可造成巨大经济损失.疫苗免疫是有效控制牛感染本病的途径.目前,S19和RB51减毒活疫苗已经被广泛用于牛布鲁氏菌病的预防.然而,如果将这些疫苗用于怀孕动物,可能会引起流产.因此,这些疫苗对于动物而言存在一定毒力,限制了其使用.为了解决这些问题,采用相关毒力基因缺失技术改善现有疫苗.
GntR家族是一类非常重要的毒力调控子,广泛分布在病原菌中,它们调控细菌的许多生物过程.GntR调控子包含从GntR1至GntR20毒力因子.质粒标记诱变技术显示羊种布鲁氏菌16MΔgntR10突变株在小鼠体内是减毒的.因此,GntR10在布鲁氏菌致病过程中扮演重要角色.本研究用2308ΔgntR10突变株感染上皮细胞系(HPT-8)、小鼠巨噬细胞系(RAW 264.7)和小鼠模型(BALB/c),目的是探索牛种布鲁氏菌2308(S2308)在细胞和小鼠模型中的胞内存活作用,揭示GntR10在细胞和小鼠感染过程中调节布鲁氏菌毒力的可能机制.
1 材料和方法
1.1 材料
1.1.1 菌株、质粒和引物
布鲁氏菌S2308、枯草芽孢杆菌(B.subtilis)为本研究室保存;自杀载体pGEM-7Zf+购自Promega公司;pMD19-T Simple克隆载体购自Takara公司;本研究所用引物及其序列见表1.

表1 本研究所用的引物
1.1.2 细胞
上皮细胞系(HPT-8)和巨噬细胞系(RAW 264.7)购自北京协和细胞资源中心.
1.1.3 小鼠
4-6周龄雌性BALB/c小鼠购自乌鲁木齐疾病预防控制中心.
1.1.4 主要试剂
2×Es Taq MasterMix、普通琼脂糖凝胶DNA回收试剂盒、高纯度质粒小提试剂盒、DNA分子量Maker、E.coli DH5α感受态细胞购自北京康为世纪生物公司;琼脂糖购自西班牙Biowest公司;限制性核酸内切酶(Sph I、Xho I、BamH I)、T4 DNA连接酶购自Takara公司;氨苄霉素购自Solarbio公司;庆大霉素购自Invitrogen公司;DMEM高糖、胎牛血清(FBS)购自Gibco公司.
1.1.5 培养基
布鲁氏菌液体培养基(TSB)和布鲁氏菌固体培养基(TSA)购自美国BD公司;布鲁氏菌鉴别培养基(BMB)、蛋白胨和酵母提取物购自OXOID公司;琼脂粉购自日本Biosharp公司;氯化钠购自上海生工生物工程股份有限公司.
1.2 方法
1.2.1 2308ΔgntR10突变株的构建
按照已公布的方法构建2308ΔgntR10突变株.以S2308基因组为模版,用GntR10-N-F和GntR10-N-R引物(表1)扩增gntR10基因的上游同源臂(957 bp).用GntR10-C-F和GntR10-C-R引物(表1)扩增gntR10基因的下游同源臂(876 bp).通过SphⅠ、XhoⅠ和BamHI位点连接至pGEM-7Zf+载体,获得重组质粒pGEM-7Zf+-gntR10.以B.subtilis为模板,用SacB-F和SacB-R引物(表1)扩增标记基因SacB(1500 bp).通过BamHI-BamHI位点,亚克隆至pGEM-7Zf+-gntR10载体,获得自杀质粒pGEM-7Zf+-gntR10-SacB.将该质粒电转化至S2308感受态细胞,涂布TSA平板(含100 μg/mL氨苄霉素),进行初次筛选.将初次筛选的细菌,涂布TSA平板(含5%蔗糖),进行第二次筛选.按照公布的方法,对第二次筛选到的细菌进行PCR鉴定.以筛选到的细菌为模板,用GntR10-F和GntR10-R引物(表1)进行鉴定,将鉴定正确的菌株命名为2308ΔgntR10.
1.2.2 互补株的构建
按照已公布的方法构建2308ΔgntR10突变株的互补株[7].以S2308基因组为模板,用GntR10-N-F和GntR10-C-R引物进行PCR扩增,将扩增到的片段连接至pMD19-T Simple载体,获得质粒GntR10-T.将该质粒电转化至2308ΔgntR10突变株内,电转后的产物涂布TSA平板(含100 μg/mL氨苄霉素).以GntR10-F和GntR10-R为引物,对互补株2308ΔgntR10-C进行RT-PCR鉴定,进一步证实gntR10基因在互补株2308ΔgntR10-C中转录恢复.
1.2.3 2308ΔgntR10在细胞模型中的毒力检测
按照已公布的方法,测定细菌在细胞模型中的毒力[8].将上皮细胞系HPT-8和巨噬细胞系RAW 264.7细胞培养至6孔板内,细胞在37 ℃、5% CO2的环境中培养至单层(约1 × 105个/孔).随后用2308ΔgntR10、2308ΔgntR10-C和S2308以100∶1的感染复数(MOI=100)侵染细胞,侵染后细胞置于37 ℃、5% CO2的培养箱内.侵染45 min后,细胞用新鲜DMEM培养液漂洗三次,随后加入含50 μg/mL庆大霉素的DMEM,孵育1 h,杀死胞外菌.1 h后弃去培养液,加入新鲜的含25 μg/mL庆大霉素的DMEM培养液(此时定义为0 h).在感染后的不同时间点(0 h、4 h、8 h、12 h、24 h和48 h),取出细胞,弃去培养液,用PBS漂洗三次,随后加入1 mL含0.1%曲拉通的PBS裂解细胞,将裂解物10倍梯度稀释后,选择合适的稀释度涂布TSA平板,对细菌进行CFU计数.实验重复3次,每次3个平行.
1.2.4 2308ΔgntR10在小鼠模型中的毒力检测
按照已公布的方法,测定细菌在小鼠模型中的毒力[9].4-6周龄雌性BALB/c小鼠,随机分成4组,分别腹腔注射200 μL 含1×106CFU的2308ΔgntR10、2308ΔgntR10-C和S2308的PBS,阴性对照组注射200 μL PBS.细菌的毒力通过测定感染后不同时间点细菌在小鼠脾脏中的数量进行确定.感染后第2、4、6和8周,各组分别处死小鼠(每组每个时间5只),无菌分离脾脏.脾脏用1 mL含0.1%曲拉通的PBS进行研磨,裂解脾细胞,将裂解物10倍梯度稀释后,选择合适的稀释度涂布TSA平板,对细菌进行CFU计数.实验重复3次,每次3个平行.
1.2.5 统计分析
细菌在细胞和小鼠模型内的存活能力用Log CFU平均值±SD表示.用SPSS 17.0分析突变株与亲本株之间的差异性,当P值< 0.05时,差异显著;P值< 0.01时,差异极显著.
2 结果与分析
2.1 2308ΔgntR10突变株和2308ΔgntR10-C互补株的构建
用GntR10-F和GntR10-R引物,可在S2308中扩增到735 bp大小的DNA片段,但是用相同的引物,在2308ΔgntR10中却没有扩增到任何片段,表明gntR10基因被敲除(图1A).此外,RT-PCR结果显示,gntR10基因在2308ΔgntR10中没有转录,但是在互补株2308ΔgntR10-C内却发生了转录,表明2308ΔgntR10-C互补株构建成功(图1B).
2.2 2308ΔgntR10在HPT-8细胞中是减毒的
布鲁氏菌侵染HPT-8细胞0 h后,各组的细菌进入胞内的数量没有差异(图2).侵染4 h后,HPT-8细胞对各组细菌的载菌量几乎相等,表明各组细菌对HPT-8细胞的黏附侵袭能力没有显著差异(图2).侵染后8 h,2308ΔgntR10突变株进入到HPT-8细胞内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降0.79-log(P> 0.05)和0.59-log(P> 0.05)(图2).侵染后12 h,2308ΔgntR10突变株进入到HPT-8细胞内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降1.08-log(P> 0.05)和0.97-log(P> 0.05)(图2).侵染后24 h,2308ΔgntR10突变株进入到HPT-8细胞内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降1.45-log(P> 0.05)和1.40-log(P> 0.05)(图2).侵染后48 h,2308ΔgntR10突变株进入到HPT-8细胞内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降1.73-log(P< 0.05)和1.54-log(P< 0.05)(图2).结果显示,与S2308相比,2308ΔgntR10突变株在HPT-8细胞中限制了细菌的繁殖能力,表明2308ΔgntR10突变株在HPT-8细胞内是减毒的.
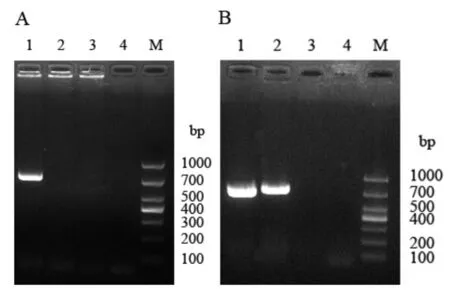
图1 2308ΔgntR10 and 2308ΔgntR10-C的鉴定

图2 2308ΔgntR10在HPT-8细胞内的胞内生存能力
2.3 2308ΔgntR10在RAW 264.7细胞中是减毒的
布鲁氏菌侵染RAW 264.7巨噬细胞0 h后,各组的细菌进入胞内的数量没有差异(图3).侵染4 h后,RAW 264.7巨噬细胞对各组细菌的载菌量几乎相等,表明各组细菌对RAW 264.7巨噬细胞的黏附侵袭能力没有显著差异(图3).然而,侵染后8 h,2308ΔgntR10突变株进入到RAW 264.7巨噬细胞内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降2.06-log(P< 0.01)和1.84-log(P< 0.01)(图3).侵染后12 h,2308ΔgntR10突变株进入到RAW 264.7巨噬细胞内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降2.78-log(P< 0.01)和2.69-log(P< 0.01)(图3).侵染后24 h,2308ΔgntR10突变株进入到RAW 264.7巨噬细胞内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降4.02-log(P< 0.01)和3.90-log(P< 0.01)(图3).侵染后48 h,2308ΔgntR10突变株进入到RAW 264.7巨噬细胞内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降4.41-log(P< 0.01)和4.29-log(P< 0.01)(图3).结果显示,与S2308相比,2308ΔgntR10突变株在RAW 264.7细胞中限制了细菌的繁殖能力,表明2308ΔgntR10突变株在RAW 264.7巨噬细胞内是减毒的.
2.4 2308ΔgntR10在BALB/c小鼠中是减毒的
布鲁氏菌感染BALB/c小鼠2周后,2308ΔgntR10突变株进入到BALB/c小鼠脾脏内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降1.52-log(P< 0.01)和1.44-log(P< 0.01)(图4).感染BALB/c小鼠4周后,2308ΔgntR10突变株进入到BALB/c小鼠脾脏内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降2.47-log(P< 0.01)和2.31-log(P< 0.01)(图4).感染BALB/c小鼠6周后,2308ΔgntR10突变株进入到BALB/c小鼠脾脏内的数量分别比亲本株S2308和互补株2308ΔgntR10-C下降2.76-log(P< 0.01)和2.63-log(P< 0.01)(图4).然而,感染BALB/c小鼠8周后,从BALB/c小鼠脾脏内仅分离到0.43-log的2308ΔgntR10突变株,分离到3.72-log的S2308,分离到3.68-log的2308ΔgntR10-C(图4).结果表明,2308ΔgntR10突变株在BALB/c小鼠体内的存活能力是降低的.

图3 2308ΔgntR10在RAW 264.7巨噬细胞内的胞内生存能力
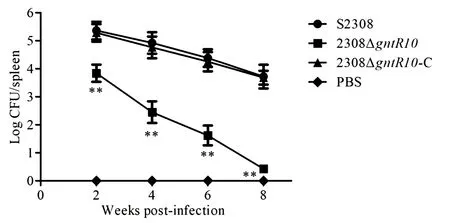
图4 2308ΔgntR10感染BALB/c小鼠后在脾脏内的清除能力
3 讨 论
转录调控因子gntR10是一个重要的毒力基因,它属于布鲁氏菌GntR家族的成员.已有研究报道,布鲁氏菌的转录调控因子GntR在控制其毒力方面扮演重要角色.布鲁氏菌是一类兼性胞内寄生菌.它可以在专业吞噬细胞如RAW 264.7巨噬细胞中生存和繁殖,也可以在非专业吞噬细胞如滋养层细胞(HPT-8)内生存.因此,我们为了确定2308ΔgntR10突变株在布鲁氏菌中的影响,用2308ΔgntR10突变株分别侵染滋养层细胞HPT-8和小鼠巨噬细胞RAW 264.7.结果发现,2308ΔgntR10突变株在滋养层细胞和巨噬细胞内的存活能力显著降低.结果表明,2308ΔgntR10突变株在细胞模型中是减毒的.
我们的前期研究证实了一些布鲁氏菌在小鼠体内存活相关的毒力基因[9].为了检测2308ΔgntR10突变株的毒力,本研究通过检测2308ΔgntR10突变株在BALB/c小鼠体内的持续存在性,进一步证实gntR10基因与布鲁氏菌的毒力有关.在本研究中,我们发现2308ΔgntR10突变株在BALB/c小鼠体内的毒力是显著降低的.结果表明,布鲁氏菌gntR10基因的缺失,导致布鲁氏菌在BALB/c小鼠体内的生存能力下降.
在本研究中,我们用自杀质粒,构建了2308ΔgntR10突变株,发现该突变株在HPT-8滋养层细胞和RAW 264.7小鼠巨噬细胞内的存活时间是缩短的.此外,该突变株在BALB/c小鼠体内的毒力也是显著下降的.然而,2308ΔgntR10突变株能否作为潜在的新型疫苗仍需进一步研究.
猜你喜欢
传染病信息(2022年6期)2023-01-12
西南农业学报(2022年5期)2022-06-06
现代畜牧科技(2021年4期)2021-07-21
中国糖料(2021年3期)2021-07-13
智慧农业导刊(2021年20期)2021-03-09
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
中国民族医药杂志(2016年9期)2016-05-09
