
互花米草入侵对东部沿海生境的影响
2020-12-21 07:24王大卫沈文星
生物学杂志 2020年6期
王大卫, 沈文星, 汪 浩
(1. 南京林业大学 经济管理学院, 南京 210037; 2. 金陵科技学院 商学院, 南京 211169)
1 研究背景和意义
草本植物互花米草(Spartinaalterniflora)已被世界上多个国家引种,并成为当今世界上最主要的生物入侵生态学和遗传学的模式植物之一[1]。我国于1991年12月正式将其从美国引入,至1993年,其在广东省台山县发展到2 000 hm2[1],随后更是占据了中国北起天津、南至广西的广大沿海滩涂,其暴发规模远大于世界上其他国家和地区,并成为我国东部沿海生物入侵的焦点问题之一。因此,2003年,我国正式将互花米草列为第一批16种外来入侵物种名单之一[2]。其中受入侵最为典型的区域为闽江河口、上海崇明东滩和江苏盐城地区等沿海地区。
分析互花米草在我国东部沿海各地入侵历史的基础,探讨其入侵的特征和现状,在分析其对生物环境影响的基础上,提出控制其入侵的防治措施,为我国今后进一步加强对互花米草的入侵管理和控制提供有效的新思路。
2 我国东部沿海生境调查及影响机理分析
2.1 东部沿海入侵现状
2.1.1 闽江口湿地入侵
氮和磷是植物成长所需蛋白质和核酸的重要组成元素,磷还可以作为限制植物生长的限制性养分[3]。布乃顺等[3]研究发现互花米草和本地植物短叶茳芏相比,入侵种互花米草分配到繁殖器官根的养分含量远高于土著种短叶茳芏植物(表1),这也是其得以成功入侵的最重要机制之一。

表1 互花米草和短叶茳芏根、茎、叶养分含量比值[3]
表1显示两者养分分配机制不同。互花米草春季的氮磷养分主要分配到叶,其次是茎,再到作为繁殖器官的根;而夏季氮则主要分配到叶,磷主要分配到根,秋冬季节则与春季相同,主要分配至叶。土著种短叶茳芏植物则与之完全不同,其各个季节的分配顺序均为从叶到茎再到根。与土著植物相比,互花米草生长过程中会利用根围区固氮生物部分缓解氮限制的压力。与此同时,其对养分的需求较低,但利用率较高,使得互花米草占生物总量比例较高[4]。由此可知,上述机理均为互花米草在闽江区入侵成功的主要因素。
2.1.2 上海崇明东滩湿地二次入侵
互花米草在崇明东滩的入侵途径主要包括有性和无性繁殖两种方式[5]。这两种入侵方式在作用和地方上各不相同。肖德荣等[6]研究发现在恢复水文的条件下,群落边缘无性繁殖是互花米草向其滩涂地区进行扩散的最主要方式,但这一方式的扩散距离有限。随后,互花米草会再利用其大量的实生苗的扩散,从远距离定居治理区域内的裸地,再逐渐形成数量庞大的群落斑块,形成二次入侵,其特征见表2。
表2显示互花米草在二次入侵过程中种子数和种子产量非常高,而地上生物量则相对较低。由此可推断出其入侵的核心是以实生苗为定值点,再通过快速、大量的无性繁殖,形成大的斑块和成片群落,助于种群的建立和维持。另一方面,有性繁殖在二次入侵过程中则主要体现在空生态位占据上,并形成了互花米草在崇明东滩地的二次入侵机制[6]。
2.1.3 盐城沿海湿地入侵
周长芳等[7]研究发现江苏省盐城市丹顶鹤保护区内互花米草与滩涂中盐蒿。芦苇滩等植物相比,在甲烷(DMS)、一氧化二氮(N2O)、二甲基硫释(CH4)、生物量和有机质的差异(见表3)影响地区温室气体排放。
表3显示互花米草无论是二甲基硫和一氧化二氮还是甲烷的排放量上均远高于其他3个区域,这充分说明了它的入侵会在很大程度上增加区域内湿地中的温室气体排放量。互花米草的有机质含量依次约为光滩和盐蒿的9.4倍和2.6倍,但仅有芦苇含量的70%左右,与3种温室气体的排放量呈正相关。由此可推测出互花米草的生物量是产生二甲基硫的主要调节因子。而互花米草迅速扩张和暴发的入侵方式,也直接导致了我国东部沿海地区温室气体排放量过高的现象[8-10]。
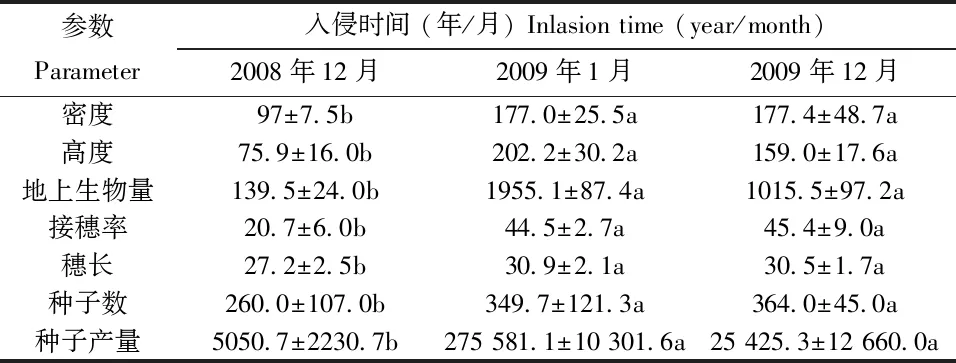
表2 互花米草二次入侵群落特征[6]
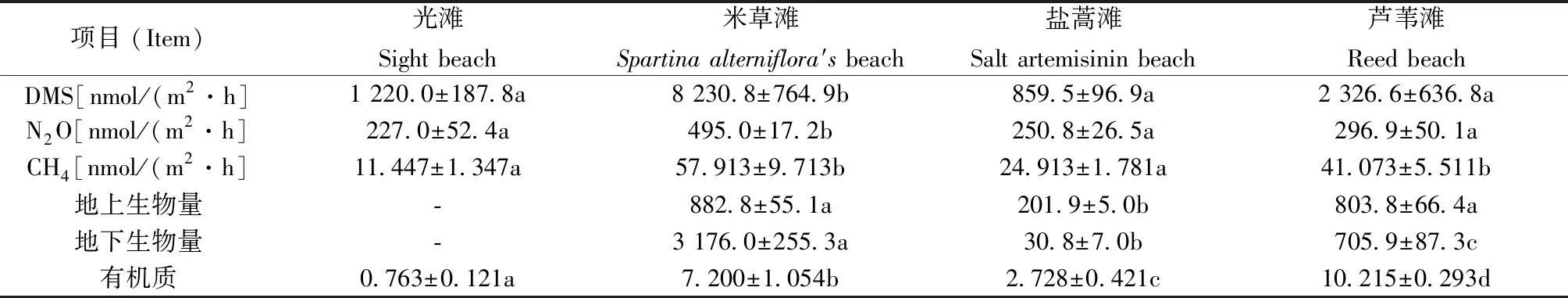
表3 不同盐沼DMS、N2O、CH4、生物量和有机质的差异[7]
2.2 互花米草的入侵机理
互花米草入侵对生态环境影响主要体现在:由于生态系统可入侵性特征,互花米草其对环境和土著生物的适应性强,以物种入侵力为基础,恰当时间的入侵通道也为成功入侵创造了入侵时机,加快了其扩张和入侵的速度[11]。同时,人类活动的影响也大大改变了生态系统的可入侵性。互花米草的入侵机制如图1所示。
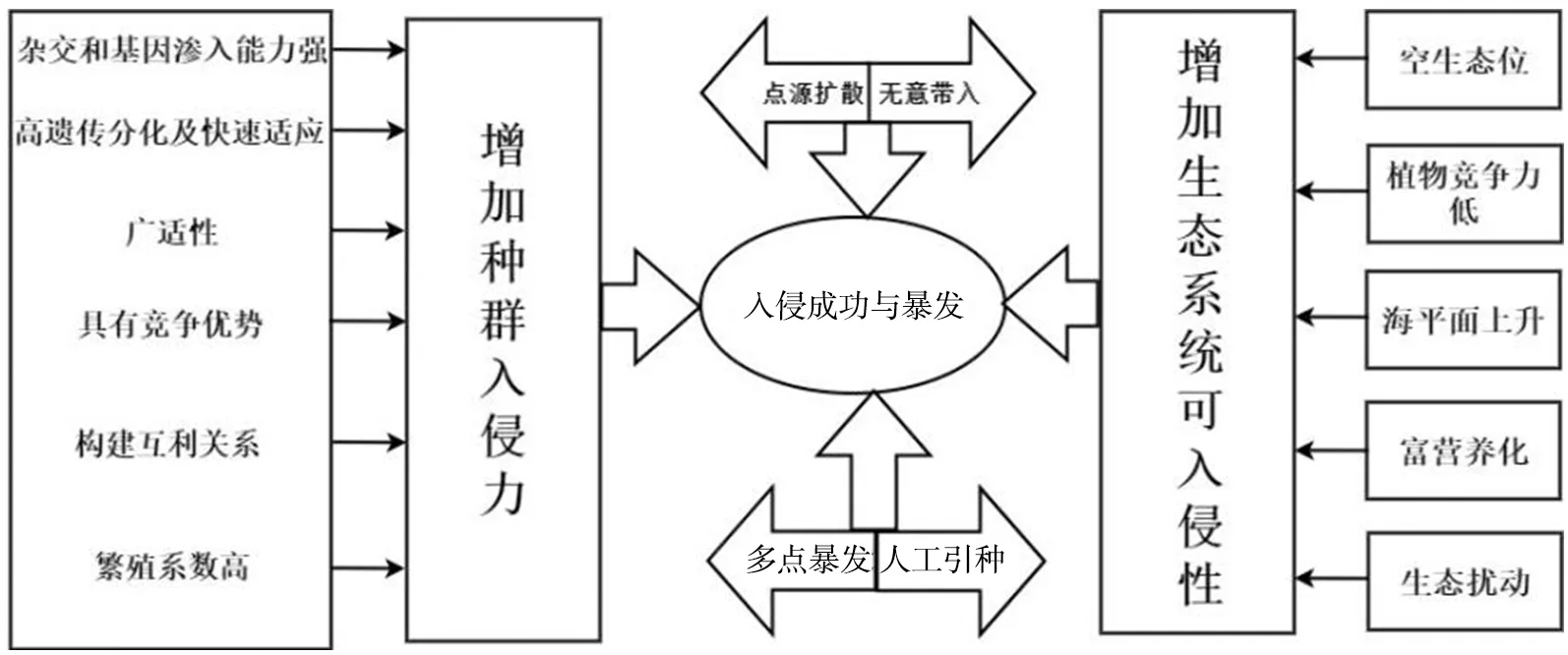
图1 互花米草种群入侵机制[11]
首先,互花米草本身的生物学特征使得其具有极强的入侵性[11]。作为其种群暴发的遗传学基础,基因渗入能力和高遗传分化使得其影响广泛。其次,作为典型的盐生植物,对盐胁迫的高抗性可使其根部更多地吸收钾离子而排斥钠离子[12]。再加上甜菜碱和脯氨酸等渗透调节物质也极大地提高了互花米草的广盐性,使得其可适应从淡水到海水的不同盐环境[13]。与此同时,互花米草的强耐淹性和高抵抗氧胁迫能力使其在被潮水淹没时的最大可耐受时间长达12 h,更进一步提高了在生态环境污染和富营养化环境里的竞争优势,增加了其入侵的机会[12-13]。
2.3 互花米草对东南沿海生境影响
互花米草对东部沿海生物环境的影响主要是对滩涂动植物和滩涂环境的影响[9]。
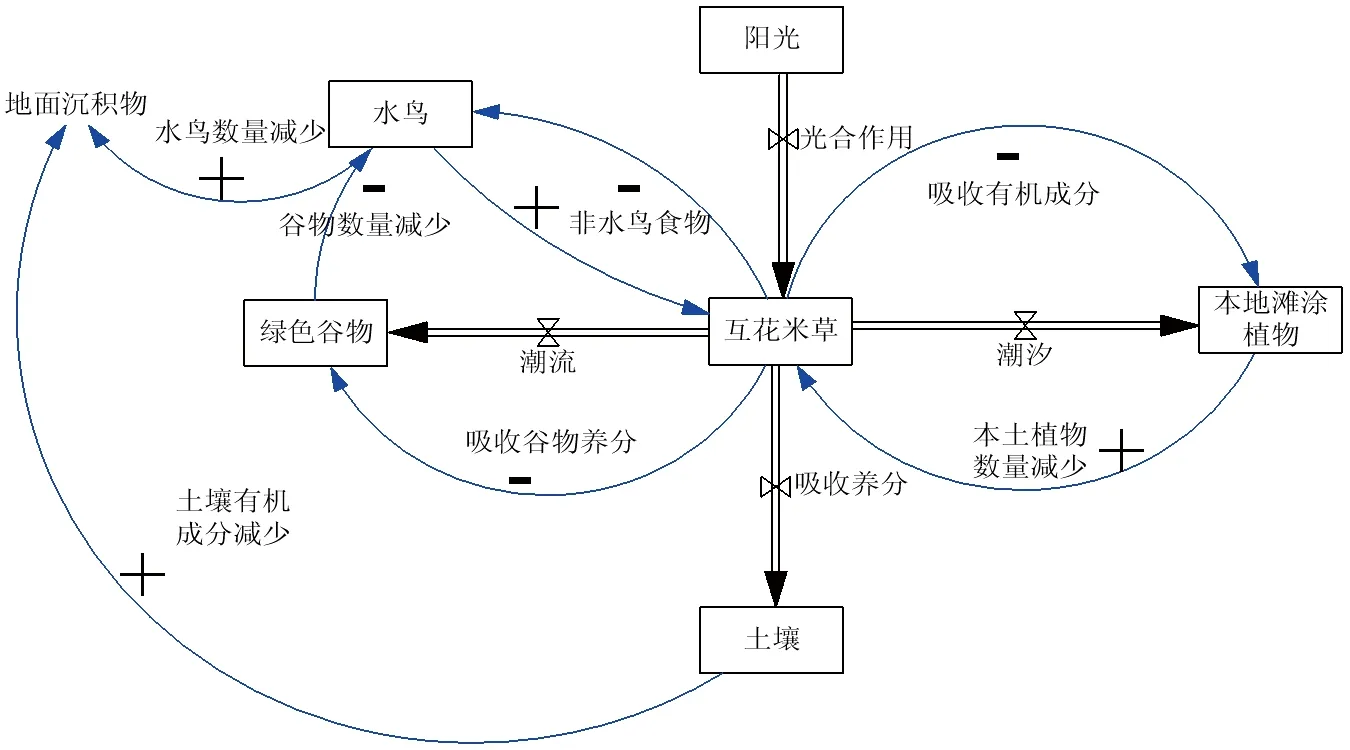
图2 互花米草对滩涂生境的影响
图2显示,在高盐度和高度淹水的环境下,互花米草利用潮汐大量吸收水流中的有机成分,从而减少当地水生植物的养分吸收以及绿色谷物的数量,由此减少本土滩涂植物数量。土壤养分也是决定物种竞争结果的重要因子,土壤养分的增加有利于外来物种的成功入侵[14]。同时,滩涂动物如水鸟等并不以互花米草作为食物,更进一步为其迅速扩张、取代本土植物、占领光滩以及形成单一密集的群落提供有利的环境。最终,鸟类生境和食物来源的丧失导致区域内水鸟种群数量急剧减少,在破坏了本土生境的生态平衡的同时,也间接导致了入侵区域的地面沉积物的大量增加,从而在很大程度上破坏了滩涂环境[15]。
2.3.1 互花米草对滩涂动物和植物的影响
互花米草的入侵往往会直接导致区域内相关动物、植物种群分布面积的大量减少以及种群数量的显著下降[16]。通过研究互花米草在我国东部海岸的入侵现状可以发现,其在入侵后会强烈排斥本土滩涂动植物,并逐渐形成显著的竞争[16]。与此同时,土壤基质和淹水等滩涂非生物因子也会促使互花米草对本土动植物的竞争关系。在盐城滨江湿地的研究发现,互花米草在有着较高的氮素水和较强的物理胁迫如高盐度和高水淹的环境下,比当地植物芦苇更有竞争优势。诸如此类的非生物因子的交互作用在很大程度上影响互花米草与本土动植物的竞争关系,进而影响了滩涂动植物种群的分布[17]。
2.3.2 互花米草对水鸟的影响
东部沿海地区的滩涂湿地是水鸟重要的栖息地与觅食地,而水鸟种群数量的维持对生态系统的稳定具有至关重要的作用。然而,入侵后的互花米草会通过占领大量吸收滩涂养分的方式排斥本土植物,抢占滩涂面积,以此造成入侵区域内生物多样性和种群的降低,其间造成大部分水鸟食物来源的缺少,由此影响水鸟种群的分布[17]。以崇明东滩为例,本土植物海三棱藨草的地下球茎和种子是白头鹤等水鸟以及一些雁鸭的主要食物来源,而互花米草在入侵后对海三棱藨草有着显著的竞争排斥,造成其种群数量短时间内大量减少。因此,当地水鸟不得不迁徙至其他地方。
综上,本地滩涂植物多样性的减少、滩涂捕食动物数量的减少以及土壤有机物中的营养减少同时增加了地面沉积物的累积,从而破坏滩涂环境。
3 互花米草的入侵控制和防治
3.1 技术防控措施
针对互花米草入侵所带来的一系列影响和问题,我国以及世界各个国家和区域均采取了有效的控制和防治措施,主要包括物理防治、化学防治、生物替代和生物防治等4种方法[19],具体如图3所示。
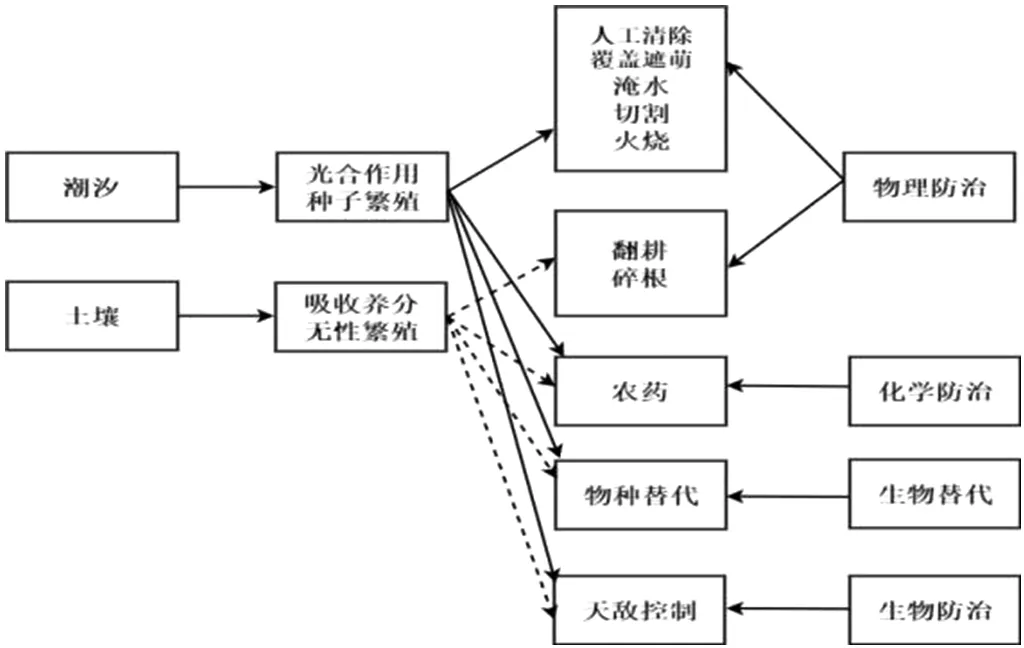
图3 互花米草控制和防治措施
物理防治措施在短期内对于较小范围内互花米草种群能够起到较为有效的抑制作用[19]。通过人工去除法,如覆盖遮阴、切割、火烧等,有效限制互花米草的呼吸和光合作用,达到杀死植株的目的[20]。化学控制措施主要通过对互花米草施用除草剂进行控制和防治工作,主要的除草剂包括草甘膦及草铵膦等。其中草甘膦是目前世界范围内应用最广、使用量最大的除草剂,但施用后的效果呈现时间较长[20]。该控制措施主要施用于我国闽江口和崇明东滩地区。生物防治的措施由来已久,主要是利用寄主范围较为专一的病原微生物和植食性动物在植物表面或植株内取食,形成虫害,将植物控制在生态、经济防治水平之下。目前该控制措施主要用于我国东部沿海盐城湿地区域。
3.2 互花米草资源化利用
我国对互花米草等生物质能源的利用形式多样,但尚处于研究阶段,研究成本较高,技术不够完善[21]。对互花米草的利用主要是对秸秆的低值化利用(如肥料化、原料化、饲料化及燃料化等利用形式),另外还包括对互花米草药用价值和耐盐基因等高新技术利用的研究。其中,有关研究表明,互花米草富含的黄酮具有抗炎、降血糖、降血脂和增强免疫力等作用。与此同时,通过提取互花米草中的功能性成分,再将其科学地添加到化妆品中可制成洗发水和护肤霜等功能性产品[22]。
4 结论与展望
互花米草在我国东部沿海地区的快速扩张和暴发给区域内的生态安全带来了严重的威胁,但至今仍缺少具有环境保护、安全、高效的统一国际性措施进行控制和防治。因此,今后的互花米草防治研究应着重加强定量化评估互花米草的入侵风险,并针对区域内的特殊环境加强综合治理技术的研究,探讨如何将入侵风险评估整合进入现有的自然保护地体系,使其更好地发挥生态保护作用[23]。与此同时,加快对互花米草的利用研究才是最行之有效的方法之一。应充分利用其药用价值或其抗盐基因培育耐盐作物等,才是今后研究的重点方向。
猜你喜欢
农业灾害研究(2022年7期)2022-09-08
安徽农业科学(2022年14期)2022-08-04
儿童故事画报·自然探秘(2022年3期)2022-04-27
幼儿教育·父母孩子版(2022年3期)2022-03-14
海洋通报(2021年5期)2021-12-21
青年文学家(2020年28期)2020-11-02
能源与环境(2020年4期)2020-09-02
少年文艺(2019年5期)2019-05-28
消费导刊(2018年10期)2018-08-20
林业与生态(2017年2期)2017-03-01
