
黄山栾树新品种‘金焰彩栾’叶片呈色的生理特性及影响因子分析
2020-12-18 03:53:58吕运舟董筱昀杨小鑫孙海楠何开跃黄利斌
植物资源与环境学报 2020年6期
吕运舟, 董筱昀, 杨小鑫, 孙海楠, 何开跃, 黄利斌,①
(1. 江苏省林业科学研究院, 江苏 南京 211153; 2. 南京林业大学生物与环境学院, 江苏 南京 210037)
随着人们对居住环境要求的不断提高,彩叶观赏植物逐渐成为现代园林配置的重要组成部分[1-2]。叶色变异植株为植物彩叶新品种选育提供了理想材料[3],并在光合作用机制[4]、激素调控[5]和功能基因组学[6]等研究领域得到广泛应用。叶片色素是叶色形成的物质基础和关键因子,多数植物叶片黄化主要是因为遗传和环境因子共同作用,影响叶绿素的合成和稳定性,从而影响不同色素间的比例[7-8]。
黄山栾树(KoelreuteriabipinnataFranch.)隶属于无患子科(Sapindaceae)栾树属(KoelreuteriaLaxm.),为高大乔木,是中国优良的乡土树种之一。该种具有多种经济价值和园林观赏价值,是园林绿化、庭院配植的理想树种[9]。‘金焰彩栾’(‘Jinyan’)为黄山栾树的一个新品种,来源于黄山栾树的天然黄化突变体,枝干呈橙黄色,秋季全株金黄,观赏价值极高[9]。迄今为止,研究者已经初步鉴定出‘金焰彩栾’体内部分表达模式与黄山栾树存在差异的色素合成基因[10-11],并对其呈色期叶片色素含量进行了初步测定和分析[12],但关于‘金焰彩栾’叶片呈色的具体生理生化机制仍不清楚,有待深入研究。
鉴于此,以黄山栾树实生苗为对照,对‘金焰彩栾’叶片发育过程中叶色参数以及叶片的色素含量和比值、含水量、细胞液pH值及可溶性糖含量的变化进行了研究,并对叶色参数与色素含量进行了多元逐步线性回归分析,以期明确影响‘金焰彩栾’叶片呈色的主要因子,为探讨其叶片呈色的生理机制及选育栾树属植物彩叶新品种奠定理论基础。
1 研究地概况和研究方法
1.1 研究地概况
实验地设在江苏省林业科学研究院苗圃地内,该苗圃地位于南京市江宁区南部,具体地理坐标为东经118°46′、北纬31°51′。该区域属北亚热带季风气候,年均温15.4 ℃,年降水量1 031 mm,无霜期227 d。土壤为黄棕壤,呈酸性(pH 5至pH 6),肥力中等。
1.2 材料
在苗圃地内分别选取3株树龄4 a、长势一致的‘金焰彩栾’嫁接苗和黄山栾树实生苗作为样株。根据‘金焰彩栾’叶片呈色的变化特征[9],分别于2015年的4月14日、4月27日、5月19日、6月16日、7月15日、8月26日、9月17日、10月28日和11月27日观察样株的叶色并拍照,同时,采集样株树梢中上部枝条上的成熟功能叶(即侧枝上第3对复叶的第3至第5枚完全展开的小叶),每株采集3枚小叶。将采集的小叶置于冰盒中带回实验室,待测。
1.3 方法
1.3.1 叶色参数及色素含量测定 参照杨淑红等[13]的方法,采用CR-410色彩色差仪(日本Konica Minolta公司)测定叶片中部(避开叶脉)的叶色参数(包括L*、a*和b*值)。参照Lichtenthaler等[14]的方法测定叶片中叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)的含量,参照Pirie等[15]的方法测定叶片中花色素苷(Acy)含量,并根据测定结果计算总叶绿素(Chl)含量(即Chla和Chlb含量之和)以及Chla/Chlb、Car/Chl和Acy/Chl值。各指标均重复测定3次。
1.3.2 含水量、细胞液pH值及可溶性糖含量测定
取新鲜叶片,先使用梅特勒-托利多AL204电子天平(精度0.1 g)称量鲜质量(FW),再置于105 ℃条件下杀青15 min,然后在75 ℃条件下烘干至恒质量,称量干质量(DW),根据称量结果计算叶片含水量(WC),计算公式为WC=〔(FW-DW)/FW〕×100%。参照马树华等[16]的方法测定叶片的细胞液pH值。参照郑恬静等[17]的方法测定叶片的可溶性糖含量。各指标均重复测定3次。
1.4 数据统计与分析
使用EXCEL 2003和SPSS 13.0软件进行数据统计和分析,采用one-way ANOVA法进行单因素方差分析,采用Duncan’s新复极差法进行多重比较。
2 结果和分析
2.1 叶色变化
观察(图1)发现,‘金焰彩栾’的新叶在3月下旬至4月下旬呈橙红色,成熟叶在5月上旬至9月下旬一直呈黄绿色,在10月上旬至11月下旬呈金黄色;而实生苗叶片在整个生长过程中基本呈绿色,仅在11月中旬开始变为黄色。
2.2 叶色相关指标变化
2.2.1 叶色参数变化 叶色参数变化统计结果(表1)表明:‘金焰彩栾’叶片的L*值在4月14日至6月16日相对稳定,自7月15日开始持续显著(P<0.05)升高,于10月28日达到最高值(82.94),随后显著降低;a*值在4月14日达到最高值(56.93),随后持续降低,在8月26日降到最低值(-21.54),随后持续回升,至11月27日达到7.81;b*值在整个实验期间呈先降低后升高的变化趋势,在5月19日降到最低值(19.69),并在11月27日达到最高值(46.35)。

A,B,C: 分别示‘金焰彩栾’在春季、夏季和秋季的叶片颜色Showing leaf color of ‘Jinyan’ in spring, summer, and autumn, respectively; D,E,F: 分别示实生苗在春季、夏季和秋季的叶片颜色Showing leaf color of seedling in spring, summer, and autumn, respectively.
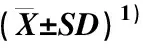
表1 黄山栾树的新品种‘金焰彩栾’及实生苗叶色参数的变化
整个实验期间,‘金焰彩栾’的L*和b*值均为正值,并且,除4月27日外,‘金焰彩栾’的L*和b*值在其余日期均极显著(P<0.01)高于实生苗。‘金焰彩栾’的a*值在4月14日、4月27日、10月28日和11月27日为正值,在其余日期为负值,而实生苗的a*值始终为负值;且‘金焰彩栾’的a*值在6月16日至9月17日低于或极显著低于实生苗,在其余日期极显著高于实生苗。
2.2.2 色素组成及比值变化 叶片色素组成及比值变化统计结果(表2)表明:整个实验期间,‘金焰彩栾’叶片叶绿素a(Chla)、叶绿素b(Chlb)和总叶绿素(Chl)含量大体呈先升高后降低的趋势,且一直维持在较低水平。‘金焰彩栾’叶片Chla、Chlb和Chl含量均在4月14日降到最低值(分别为0.05、0.01和0.06 mg·g-1),并随着时间推移而升高,在9月17日达到最高值(分别为1.95、0.28和2.23 mg·g-1),随后降低。整个实验期间,‘金焰彩栾’叶片Chla、Chlb和Chl含量基本上极显著低于实生苗。此外,整个实验期间,‘金焰彩栾’叶片Chla/Chlb值始终高于实生苗,且除11月27日外,其余日期‘金焰彩栾’和实生苗间的Chla/Chlb值均差异显著或极显著。
整个实验期间,‘金焰彩栾’叶片类胡萝卜素(Car)和花色素苷(Acy)含量均呈明显的波动变化,分别在4月27日和10月28日达到最高值,在11月27日分别为4月14日的3.4和1.3倍;而‘金焰彩栾’叶片Car/Chl和Acy/Chl值则大体呈“迅速降低—保持稳定—逐渐升高”的趋势。另外,整个实验期间,‘金焰彩栾’叶片Car和Acy含量极显著低于实生苗,而叶片Car/Chl和Acy/Chl值却显著或极显著高于实生苗(6月16日除外)。
表2 黄山栾树的新品种‘金焰彩栾’及实生苗叶片色素含量及比值的变化
2.2.3 含水量、细胞液pH值和可溶性糖含量变化
统计结果(表3)表明:整个实验期间,‘金焰彩栾’叶片含水量大体呈逐渐降低的趋势;细胞液pH值大体呈先降低后升高的趋势;可溶性糖含量大体呈“降低—升高—降低—升高”的趋势,在4月27日降到最低值(10.63 mg·g-1),在7月15日达到最高值(24.89 mg·g-1)。
除11月27日外,其余日期‘金焰彩栾’和实生苗间的叶片含水量均差异显著或极显著,且除8月26日外,‘金焰彩栾’叶片含水量均高于实生苗。‘金焰彩栾’叶片细胞液pH值在4月27日极显著高于实生苗,在5月19日和7月15日极显著低于实生苗,在9月17日和10月28日显著低于实生苗。‘金焰彩栾’叶片可溶性糖含量在4月14日和11月27日均极显著高于实生苗,在8月26日极显著低于实生苗,在6月16日和9月17日显著低于实生苗。
表3 黄山栾树的新品种‘金焰彩栾’及实生苗叶片含水量、细胞液pH值和可溶性糖含量的变化
2.3 多元逐步线性回归分析
分别以L*、a*和b*值为因变量,‘金焰彩栾’叶片总叶绿素(Chl)、类胡萝卜素(Car)和花色素苷(Acy)含量为自变量,进行多元逐步线性回归分析,获得的回归方程分别为L*=32.776+100.583Car(R=0.619,P<0.05)、a*=38.829-30.399Chl(R=0.724,P<0.05)和b*=9.414+78.124Car(R=0.533,P<0.05),说明‘金焰彩栾’叶片的叶色参数与Chl和Car含量密切相关。其中,a*值与Chl含量显著负相关,即随着Chl含量增加,叶片绿色调增强;L*和b*值与Car含量显著正相关,即随着Car含量增加,叶片黄色调和整体明度增强。
3 讨论和结论
研究表明:色素类型和含量是影响植物叶色的最主要和最直接的因子[18-20],多数植物叶色突变体表型由叶绿素含量下降引起[21-23]。本研究结果也表明:在整个实验期间,黄山栾树新品种‘金焰彩栾’叶片的叶绿素a(Chla)、叶绿素b(Chlb)和总叶绿素(Chl)含量基本上极显著低于实生苗,而‘金焰彩栾’叶片的Chla/Chlb值却基本上显著或极显著高于实生苗,说明‘金焰彩栾’叶片的叶绿素合成通路可能受阻,且Chlb含量的降幅更大。然而,关于‘金焰彩栾’叶片叶绿素合成受阻及调控通路的分子机制尚不清楚,有待进一步深入研究。叶绿素a酸酯氧化酶(chlorophyllideaoxygenase,CAO)是催化Chla转化成Chlb的关键酶[24],‘金焰彩栾’叶片CAO基因的表达水平显著低于实生苗[10],说明‘金焰彩栾’可通过调控CAO基因的转录表达水平调控Chlb的合成,从而提高Chla/Chlb值,使叶片呈现黄色。另外,多元逐步线性回归分析结果表明:‘金焰彩栾’叶片a*值与Chl含量显著负相关,L*和b*值与Car含量显著正相关,说明‘金焰彩栾’叶片呈色与Chl和Car含量密切相关。
郁万文等[25]的研究结果表明:Car/Chl值升高是银杏(GinkgobilobaLinn.)金叶品种‘万年金’(‘Wannianjin’)半同胞子代无性系叶片呈黄色的主要原因。本研究中,实验开始时(即4月14日),‘金焰彩栾’叶片的Chl含量极低(0.06 mg·g-1),但Car/Chl和Acy/Chl值却最大,叶片呈橙红色;随后,Car/Chl值持续下降,当Car/Chl值低于0.28时,叶片呈黄绿色或绿色;入秋后,Chl含量下降,但Car/Chl值升高,叶片呈金黄色。说明‘金焰彩栾’叶片呈色与Car/Chl值有关。
综上所述,‘金焰彩栾’叶色的适宜观赏期为春季和秋季,夏季叶片出现复绿现象,观赏价值降低。‘金焰彩栾’叶色变化是多种因子综合作用的结果,Chl和Car含量及Car/Chl值为主要影响因子。
猜你喜欢
热带作物学报(2023年7期)2023-08-14 00:07:07
农业科技通讯(2022年1期)2022-01-17 06:06:40
天天爱科学(2020年11期)2020-09-10 17:17:53
山东林业科技(2019年2期)2019-06-03 10:10:54
作文成功之路·教育教学研究(2019年2期)2019-03-20 12:34:32
作文成功之路·小学版(2019年2期)2019-01-14 16:41:44
——以新源县为例
新疆农业科技(2018年5期)2018-05-07 02:25:08
课程教育研究(2017年20期)2017-06-14 11:41:41
长江文艺(2017年2期)2017-02-13 19:17:24
中国糖料(2016年1期)2016-12-01 06:48:57
