
外源激素对低温胁迫下南美油藤幼苗叶片生理生化指标的影响
2020-12-11 09:10刘果高丽琼彭彦尚秀华詹妮吴志华
南方农业学报 2020年9期
关键词:低温胁迫
刘果 高丽琼 彭彦 尚秀华 詹妮 吴志华

摘要:【目的】探索南美油藤幼苗的抗冷害機制,为我国南美油藤幼苗低温胁迫适应性研究提供一定的理论参考和支持。【方法】利用脱落酸(ABA)和赤霉素(GA3)两种外源激素作为诱导剂,测定南美油藤幼苗在4 ℃低温胁迫下不同激素和不同浓度处理下的幼苗叶片膜系统[相对电导率(REC),丙二醛(MDA)含量]、渗透调节物质[可溶性蛋白(SP),可溶性糖(SS)]和氧化酶[超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)]活性等生理生化指标随处理时间的变化。【结果】南美油藤幼苗叶片喷施10 μmol/L ABA后经低温胁迫能显著降低其膜系统REC和MDA含量(P<0.05,下同),而喷施10 mg/L GA3能有效降低其MDA含量;南美油藤幼苗叶片喷施10 μmol/L ABA后经低温胁迫能显著增加其SP和SS含量,而喷施GA3南美油藤幼苗叶片SP和SS含量的增加效果不显著(P>0.05,下同);喷施10 μmol/L ABA后经低温胁迫均能有效提高幼苗叶片SOD、POD和CAT活性,喷施10 mg/L GA3可适当提高其POD活性,5 mg/L GA3能在一定程度上提高其CAT活性,但随着低温处理时间的延长,喷施GA3对南美油藤幼苗叶片中CAT活性增强效果不显著。对7种生理生化指标的Pearson相关性分析结果显示,SOD、POD和CAT 3种氧化酶活性间存在正相关关系,且POD与REC呈负相关,CAT与REC和MDA含量呈负相关,故增强SOD、POD和CAT的活性,可适当降低REC和MDA含量,提高南美油藤幼苗的抗冷害能力。SP和SS均与3种氧化酶活性呈正相关,且SS与REC和MDA呈负相关,因此增加SP和SS含量对南美油藤幼苗抵抗冷害能力有积极的作用。【结论】喷施适当浓度的外源激素ABA(10 μmol/L)和GA3(5~10 mg/L)能有效提高南美油藤幼苗的抗冷害能力。
关键词: 南美油藤;外源激素;低温胁迫;氧化酶活性
中图分类号: S565.9 文献标志码: A 文章编号:2095-1191(2020)09-2181-10
Effects of exogenous hormones on physiological and biochemical indexes of Plukenetia volubilis under low temperature stress
LIU Guo, GAO Li-qiong, PENG Yan, SHANG Xiu-hua, ZHAN Ni, WU Zhi-hua*
(Center for Eucalyptus Research and Development, National Forestry and Grassland Administration,
Zhanjiang, Guangdong 524022, China)
Abstract:【Objective】Mechanism of Plukenetia volubilis seedlings resistance to cold damage was explored to provide reference for adaptation analysis of P. volubilis seedlings responding to low temperature stress. 【Method】In this study, two exogenous hormones, abscisic acid(ABA) and gibberellin(GA3), were used as inducers to determine the changes of physiological and biochemical indicators such as blade membrane system[relative conductivity(REC), the content of malonaldehyde(MDA)], osmotic adjustment substances[soluble protein(SP) and soluble sugar(SS)], oxidase[activity of superoxide dismutase(SOD), peroxidase(POD) and catalase(CAT)] in the leaves of P. volubilis seedlings treated with different hormones concentrations and the treatment times under low temperature stress. 【Result】The results showed that 10 μmol/L ABA applied to the leaf membrane system of seedlings after low temperature stress could significantly reduce their REC and MDA content(P<0.05, the same below), and 10 mg/L GA3 could effectively reduce the content of MDA. 10 μmol/L ABA could significantly increased SP and SS content in P. volubilis seedlings osmotic adjustment substance, but the effect of GA3 was not significant in increasing the content of SP and SS under low temperature stress(P>0.05, the same below). Additionally, 10 μmol/L ABA could effectively improve SOD, POD and CAT activities of seedlings leaves, 10 mg/L GA3 could properly improve POD activity and 5 mg/L GA3 could improve CAT activity to some extent, but the effect of GA3 on enhancement of CAT activity was not significant with the extension of time. The Pearson correlation analysis was carried out on the seven physiological and biochemical indexes. The results showed that there were positive correlation relationships among SOD, POD and CAT, and POD was negatively correlated to REC, CAT were negatively correlated to REC and MDA. Therefore, the increase of SOD, POD and CAT activities could decrease the REC and the content of MDA which could improve the ability of cold resistant of P. volubilis seedlings. SP and SS were positively correlated with the activities of SOD, POD and CAT, and SS was negatively correlated with REC and MDA, thus increasing the contents of SP and SS could have a positive effect on the cold resistant of P. volubilis seedlings. 【Conclusion】Spraying with appropriate concentration of exogenous hormone ABA(10 μmol/L) and GA3(5-10 mg/L) could effectively enhance the ability of cold resistant of P. volubilis seedlings.
Key words: Plukenetia volubilis; exogenous hormones; cold resistant; the oxidase activity
Foundation item:Basic and Applied Basic Research Project of Guangdong(2019A1515110168); Forestry Science and Technology Innovation Project of Guangdong(2017KJCX015)
0 引言
【研究意义】南美油藤(Plukenetia volubilis)是大戟科攀援状多年生木质藤本植物,又名印加果、星油藤、美藤果等,原生长在海拔80~1700 m的南美洲安第斯山脉地区热带雨林(de Souza et al.,2013)。南美油藤种子含油量丰富(35%~60%)(Kumar et al.,2017),种子油的不饱和脂肪酸含量高达90%以上,且不含任何毒素和对健康有害的物质,是世界上不饱和脂肪酸含量最高的植物油之一(Chandrase-karan and Liu,2015);同时,南美油藤种子油组分优良,营养品质高,被认为是世界上最好的植物油之一(刘果等,2018)。我国自2006年首次引种以来,已在云南南部、贵州、海南和广东等地区有少量分布。南美油藤原生长地为热带或亚热带型气候,温度为10~26 ℃,相对湿度70%~80%(Alicia and Isabella,2016)。与原产地相比较发现,我国目前南美油藤的适生区普遍存在冬季气温低于10 ℃的情况,因此,提高南美油藤植株,特别是幼苗的抗冷害能力对发展南美油藤产业具有非常重要的意义。【前人研究进展】温度是重要的环境因子之一,在限制植物遗传背景的前提下,温度对植物生长发育的某些过程起决定性作用(闫绍鹏,2004)。在低温条件下,植物体内会产生各项生理生化的变化,如生物膜的渗透性、结构和组分、细胞质的流动性、酶活性以及呼吸作用和光合作用等的變化,以降低植物受低温伤害的程度(苏鹏,2010)。作为抗寒冷基因表达的启动因素之一,植物激素在植物适应生物和非生物胁迫中起着积极的调控作用(罗正荣,1989)。研究表明,脱落酸(Abscisic acid,ABA)和赤霉素(Gibberellin,GA3)的含量与植物的抗寒性密切相关(王连荣等,2016)。已有研究表明,甘蔗(Saccharum officenarum)(黄杏,2012)、杏扁(Prunus armeniace)(王连荣等,2016)、大豆(Glycine max)(魏鑫等,2016)、玉米(Maize)(李馨园等,2017)等外施适宜浓度的ABA后,可在不抑制生长的情况下提高植株抗寒力。GA3是植物生长和发育过程中必不可少的激素之一,其参与调控植物种子萌发、叶片生长及植物开花等生长发育的各个方面(Richards et al.,2001)。代勋等(2012)研究表明,喷施10 mg/L GA3可提高低温胁迫下小桐子(Jatropha curcas)种子的发芽率,并能显著提高幼苗对低温胁迫的抵抗能力。Zhu等(2016)的研究显示,经0.5-mM GA3处理后,能有效抑制番茄(Solanum lycopersicum)果实冷害指数的上升。【本研究切入点】南美油藤的产业化发展和应用前景广阔,但目前在我国的分布区域有限,仅华南和西南部分地区有少量种植。近年来,关于南美油藤的研究多集中于脂肪酸生物合成与累积的机制探析(Chandrasekaran and Liu,2015; Hu et al.,2018; 刘果等,2019),针对低温胁迫下南美油藤幼苗各项生理生化特性的研究鲜有报道。【拟解决的关键问题】以当年生南美油藤幼苗为试验材料,分别研究外源ABA和GA3对低温处理后南美油藤幼苗对低温胁迫的响应,以期寻找最优的浓度,为我国南美油藤幼苗低温胁迫适应性研究提供一定的理论参考和支持。
1 材料与方法
1. 1 试验材料
试验用南美油藤果于2019年2月采自广东省湛江市遂溪县南方国家级林木种苗示范基地内南美油藤种质资源圃。果实采摘后自然晾干,剥壳获得的健康种子经灭菌消毒后点播在装满轻型基质(黄心土∶稻壳∶珍珠岩=3∶4∶3)的育苗袋(12 cm×14 cm)中,每个育苗袋点播1颗种子。种子萌发后,于2019年4月初选取生长健壮且形态相对一致的南美油藤幼苗置于南方国家级林木种苗示范基地高新温室内进行正常生长培育。
1. 2 试验方法
南美油藤正常生长3个月后开始进行处理。参考罗银玲等(2014)的方法,分别用500 mL水(对照,CK)及不同浓度的ABA溶液(5、10和50 μmol/L,分别标记为ABA-1,ABA-2和ABA-3)和GA3溶液(5、10和50 mg/L,分别标记为GA-1,GA-2和GA-3)对南美油藤幼苗进行叶面喷施,每处理30株幼苗。早上9:00第1次喷施后,于次日9:00进行第2次叶面喷施。第3 d早上9:00将幼苗置于4 ℃光照培养箱(光照强度为800 lx)中进行冷害处理,分别于低温处理24、48和72 h后取6株幼苗的第3对叶子,将所有叶片保存在-20 ℃冰箱中用于测定各种生理生化指标。以喷施水但未进行低温处理的幼苗为对照1(CK-1),喷施水且进行低温处理的幼苗记为对照2(CK-2)。
1. 3 测定项目及方法
相对电导率(Relative electric conductivity,REC)采用电导法测定;丙二醛(Malondialdehyde,MDA)含量采用硫代巴比妥酸酸(TBA)法测定;可溶性蛋白(Soluble protein,SP)含量采用考马斯亮蓝法测定;可溶性糖(Soluble sugar,SS)含量采用蒽酮法测定;超氧化物歧化酶(Superoxide dismutase,SOD)活性采用氮蓝四唑(NBT)显色法测定;过氧化物酶(Peroxidase,POD)活性采用愈创木酚比色法测定。过氧化氢酶(Catalase,CAT)活性的测定方法为:取0.15 mol/L pH 7.0的磷酸缓冲液200 mL,加入0.309 mL H2O2(原液)摇匀即为反应液;3 mL反应液中加入0.1 mL上述提取的酶液,以pH 7.0的磷酸缓冲液对照调零,在240 nm波长下测定吸光度值。
1. 4 统计分析
试验数据运用Excel 2016进行作图分析,并结合SPSS 19.0进行方差分析、Pearson相关性分析和t检验的差异分析。
2 结果与分析
2. 1 外源激素对低温胁迫下南美油藤幼苗叶片膜系统的影响
2. 1. 1 REC 根据t检验结果(表1)可知,经低温处理后,各处理组的南美油藤幼苗叶片REC与CK-1达显著(P<0.05,下同)或极显著(P<0.01,下同)差异水平,其中ABA-1、GA-1、GA-2和GA-3处理组的幼苗叶片REC与CK-1呈极显著差异。ABA-2处理组的幼苗叶片REC与除ABA-3处理组外的其他处理组均存在显著差异,此外其与除CK-2外的其他处理组间差异不显著(P>0.05,下同),由此表明,ABA-2处理组幼苗叶片REC较其他处理组的变化更小,幼苗叶片的细胞内液更稳定。
从图1可知,经低温处理后,南美油藤幼苗叶片REC显著升高。未喷施激素的CK-2受低温伤害明显,其REC在低温处理48 h时达最大值。4 ℃低温处理72 h后,进行激素喷施的处理中,ABA-2处理组的幼苗叶片REC较其他处理低,且ABA处理组整体REC低于GA3处理组,即幼苗叶片的细胞内液渗出更少,能有效降低低温对南美油藤幼苗的伤害,说明ABA对降低REC的作用较GA3更好。从图1还可知,随着处理时间的延长,低温胁迫下的南美油藤幼苗叶片REC以ABA-2处理组变化较为平缓,且与其他处理差异显著;ABA-1和GA-3处理组的幼苗叶片REC随着低温处理的时间增加而逐渐增加,而其他处理组均在48 h达最大值,处理72 h的REC值反而降低。由此推断,喷施适当浓度ABA和GA3能在一定程度上提高南美油藤幼苗的低温适应能力,以ABA-2处理组对南美油藤幼苗叶片REC的降低更显著,即ABA喷施浓度为10 μmol/L时效果最佳。
2. 1. 2 MDA含量 由图2可知,4 ℃低温胁迫下,CK-2的叶片MDA含量随着处理时间的延长而逐渐增加,其幼苗叶片受到低温胁迫的不断威胁。经激素处理的幼苗叶片MDA含量均较CK-2低,且随着处理时间的延长,ABA-2处理组的幼苗叶片MDA含量呈下降趋势,由此说明,喷施ABA浓度为10 μmol/L能有效缓解南美油藤叶片MDA含量的增加。经过72 h的低温处理,ABA-3和GA-2处理组的幼苗叶片MDA含量较低,表明ABA浓度为50 μmol/L或GA3浓度为10 mg/L时,南美油藤幼苗叶片MDA含量表现较低。根据MDA含量与ABA浓度的变化可知,喷施一定浓度的ABA和GA3可降低叶片MDA含量,其中,ABA的浓度越高,叶片MDA含量越低,即在适宜的范围内,提高ABA的浓度能有效降低MDA含量,提高南美油藤幼苗的抗冷害能力。
根据t检验结果(表2)可得,ABA-2处理组和CK-2与GA-2处理组的MDA含量均存在显著差异,而其他各处理组间的幼苗叶片MDA含量均差異不显著。由此可推断,喷施浓度为10 mg/L GA3更能有效降低低温胁迫下南美油藤幼苗叶片的MDA含量。
2. 2 外源激素对低温胁迫下南美油藤幼苗叶片渗透调节物质的影响
2. 2. 1 SP含量 从图3可知,ABA喷施浓度为10 μmol/L(ABA-2)时,南美油藤幼苗叶片SP含量在低温处理48和72 h均为最高,且高于其他处理组;而ABA浓度较低(处理24 h除外)或较高时,低温胁迫下南美油藤幼苗叶片SP含量与CK-2差异不显著。喷施不同浓度GA3的南美油藤幼苗叶片SP含量的变化与CK-2相比,GA-1和GA-2处理组的南美油藤幼苗叶片SP含量在低温处理48 h时略高于CK-2,但差异均不显著。由此表明,喷施适宜浓度的ABA(10 μmol/L)能显著提高南美油藤幼苗叶片SP含量,增强其抗低温能力;但喷施GA3对南美油藤幼苗叶片SP含量的影响不大。
从不同处理时间对南美油藤幼苗叶片SP含量的影响(图3)还发现,ABA-2、GA-1和GA-2处理组幼苗在受到低温胁迫48 h时,叶片SP含量均有显著升高,其中以ABA-2处理组幼苗叶片SP含量增加更为显著。由此可知,喷施10 μmol/L ABA能显著提高受到低温胁迫的南美油藤幼苗叶片SP含量,喷施较低浓度(5和10 mg/L)GA3对低温胁迫下南美油藤幼苗叶片SP含量稍有提高,但效果不显著。结合差异分析结果(表3)可得,喷施GA3在提高低温胁迫下南美油藤幼苗叶片SP含量的效果有限。
2. 2. 2 SS含量 从图4可知,经ABA和GA3处理的南美油藤幼苗叶片SS含量有所增加,其中以喷施10 μmol/L ABA处理组的南美油藤幼苗叶片SS含量最高。随着低温处理时间的延长,GA-1和GA-3处理组幼苗叶片SS含量逐渐降低;GA-2处理组幼苗叶片SS含量在低温处理48 h时达最大值,但在低温处理72 h后急剧降低。由此可知,GA3对低温胁迫下南美油藤幼苗叶片SS含量的增加有一定的促进作用,但随着低温时间的延长,其促进作用有限。
从图4还可知,低温处理24 h,各激素处理组和CK-2的南美油藤幼苗叶片SS含量均高于CK-1,说明南美油藤幼苗受到了低温胁迫的危害;而ABA-1、ABA-2、ABA-3和GA-1处理组的幼苗叶片SS含量均高于CK-2,由此说明,4 ℃低温胁迫下上述4个处理组的幼苗叶片SS含量增加,抵抗冷害能力增加。低温处理48 h,各激素处理组的幼苗叶片SS含量均显著高于CK-2,由此说明喷施ABA和GA3均能在一定程度上增加南美油藤幼苗叶片SS含量,且喷施ABA的效果更显著。低温处理72 h,CK-2的幼苗叶片SS含量稍有增加,说明经过72 h的低温胁迫,南美油藤幼苗叶片开始出现低温适应能力的提高,同时,除ABA-1外各处理组的幼苗叶片SS含量均呈下降趋势,仍以ABA-2处理组幼苗叶片SS含量最高,而喷施GA3各处理组的幼苗叶片SS含量与CK-2基本一致,由此说明,喷施GA3提高低温胁迫下南美油藤幼苗叶片SS含量的效果有限,结合t检验结果(表4)可得出,喷施浓度为10 μmol/L ABA能有效增加低温胁迫下南美油藤幼苗叶片SS含量。
2. 3 外源激素对低温胁迫下南美油藤幼苗叶片氧化酶活性的影响
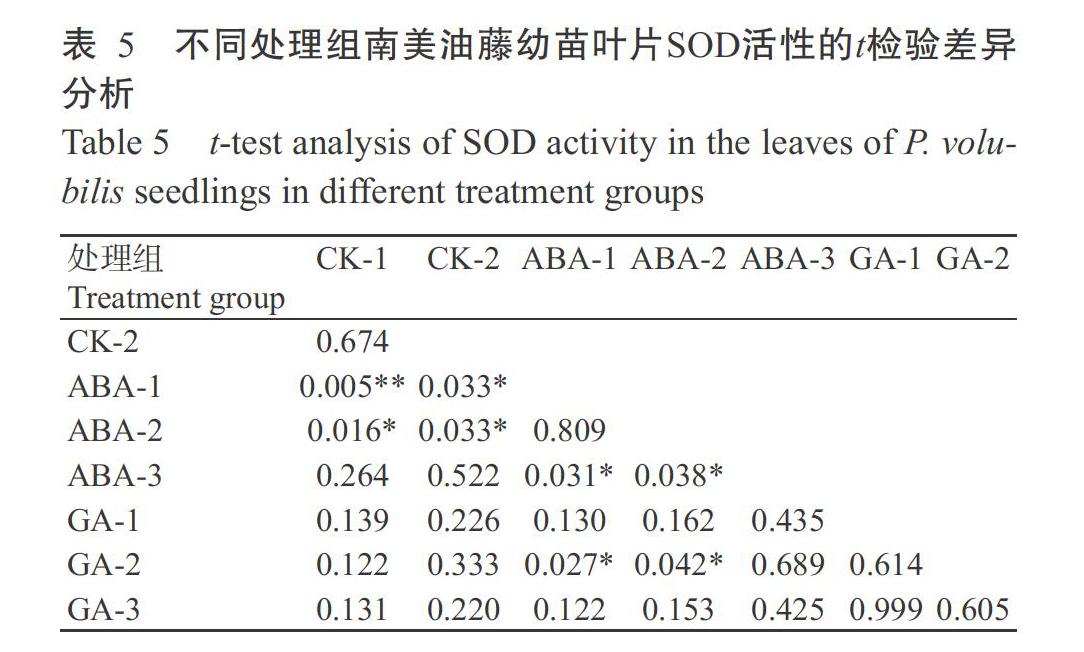
2. 3. 1 SOD活性 从图5可知,未喷施激素且经低温处理(CK-2)的南美油藤幼苗叶片SOD活性在低温处理24 h后显著降低,其余各处理组的南美油藤幼苗叶片SOD活性显著增加。低温胁迫48 h后,CK-2的幼苗叶片SOD活性急剧增加,而在低温处理72 h后又急剧降低,由此可推断,南美油藤幼苗叶片已受到低温的危害。ABA-3、GA-1、GA-2和GA-3处理组在低温处理72 h,其幼苗叶片SOD活性降低,且与CK-2的幼苗叶片SOD活性基本一致,由此可知,随着低温胁迫时间的延长,喷施高浓度ABA(ABA-3处理组)和喷施GA3处理的南美油藤幼苗叶片SOD活性与未喷施激素处理的基本一致。ABA-1和ABA-2处理组的幼苗叶片SOD活性较其他处理组显著增加,由此可知,喷施ABA浓度为5和10 μmol/L时可显著提高低温胁迫下南美油藤幼苗叶片SOD活性。
由图5还可知,经ABA和GA3喷施处理后,南美油藤幼苗叶片SOD活性显著增强。其中以ABA-1和ABA-2处理组的幼苗叶片SOD活性在不同处理时间的活性较高。根据t检验结果(表5)可得,ABA-1和ABA-2处理组的南美油藤幼苗叶片SOD活性与CK-2差异显著,由此可得,喷施ABA浓度为5和10 μmol/L时能显著提高南美油藤幼苗在低温胁迫下叶片SOD活性。喷施GA3的各处理组中,GA-3处理组的幼苗叶片SOD活性较其他两个处理组略高,但与CK-2的幼苗叶片SOD活性差异不显著。由此推断,喷施GA3能增加低温胁迫下南美油藤幼苗叶片SOD活性,但效果不显著。
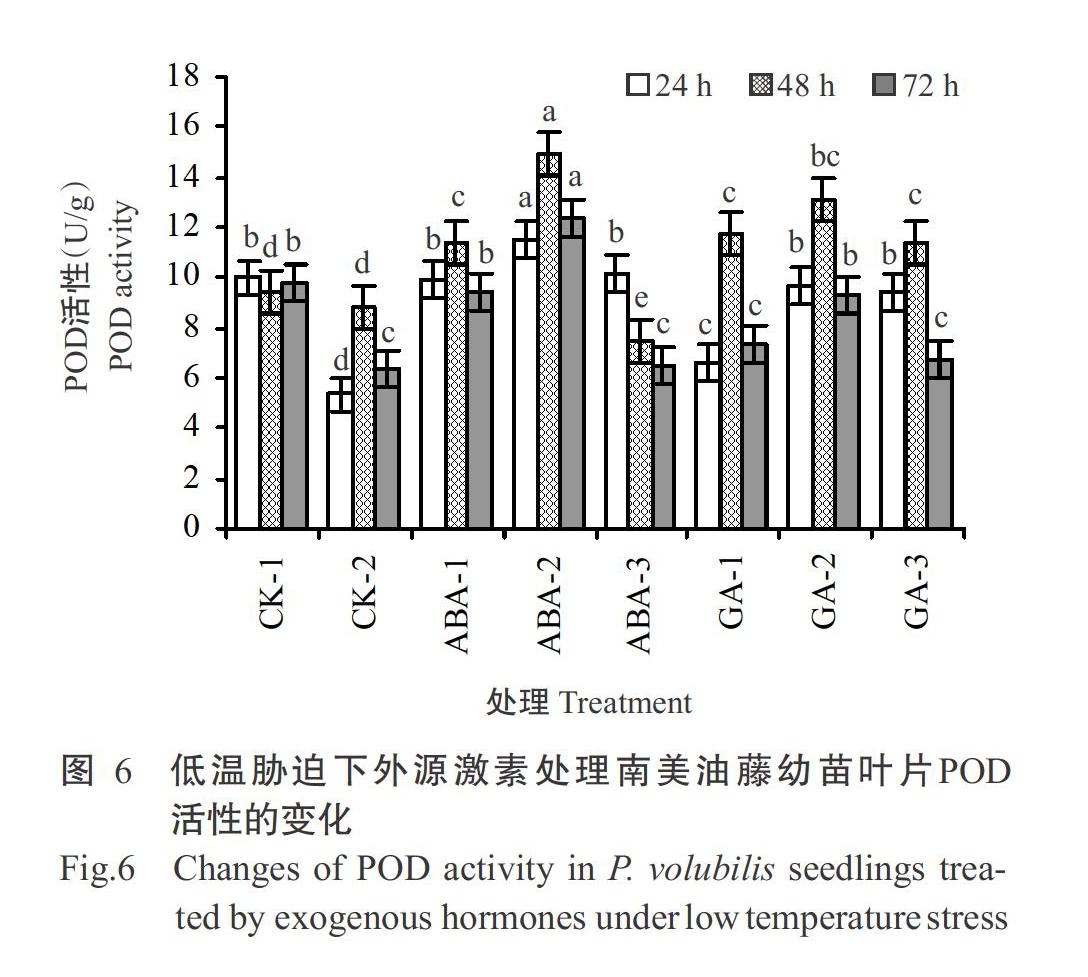
2. 3. 2 POD活性 与低温胁迫下未喷施激素处理的CK-2相比较,经ABA和GA3喷施处理后的南美油藤幼苗叶片POD活性变化存在较大差异(图6)。其中,低温处理24 h时,ABA-2处理组的幼苗叶片POD活性最高,CK-2的幼苗叶片POD活性最低;低温处理48 h时,ABA-1、ABA-2、GA-1、GA-2和GA-3处理组的幼苗叶片POD活性均明显升高,以ABA-2处理组的幼苗叶片POD活性最高;低温处理72 h时,仍以ABA-2处理组幼苗叶片POD活性最高,且与其他处理组差异显著,而喷施GA3所有处理组的幼苗叶片POD活性均有所降低。由此可推断,喷施较低浓度ABA(5和10 μmol/L)或较高浓度GA3(10和50 mg/L)能有效提高低温胁迫下南美油藤幼苗叶片POD活性;随着低温胁迫时间的延长,喷施ABA和GA3的南美油藤幼苗叶片POD活性随之降低。
根据t检验结果(表6)可知,ABA-2处理组与未喷施激素低温处理CK-2的幼苗叶片POD活性存在显著差异。由此可知,喷施ABA浓度为10 μmol/L时,对低温胁迫下南美油藤幼苗叶片POD活性增强有显著效果,喷施GA3的各处理组与CK-2的幼苗葉片POD活性均未达到显著差异水平,因此,外源喷施GA3对低温胁迫下南美油藤幼苗叶片POD活性增强效果不显著。从表6可知,GA-2处理组与CK-2的P=0.074,结合喷施GA3能在一定程度上增加幼苗叶片POD活性,且GA-2处理组POD活性增加最为显著,由此可推断,喷施GA3浓度为10 mg/L时,可在一定程度上提高低温胁迫下南美油藤幼苗叶片POD活性。
2. 3. 3 CAT活性 由图7可知,不同低温胁迫时间下各处理组的南美油藤幼苗叶片CAT活性存在差异,其中,以GA-1处理组南美油藤幼苗叶片CAT活性在处理24 h时最高,且与其他处理组差异显著;ABA-2处理组幼苗叶片在低温胁迫下48和72 h时CAT活性均最高,且与其他处理组差异显著。由此可推断,经ABA和GA3喷施处理后,南美油藤幼苗叶片CAT活性增强,可在一定程度上增加南美油藤幼苗抵抗冷害的能力,以ABA-2处理组幼苗叶片CAT活性增强最稳定。
由t检验结果(表7)可知,ABA-2处理组与CK-2呈极显著差异,说明在低温胁迫下,喷施ABA浓度为10 μmol/L时南美油藤幼苗叶片CAT活性的提高达到显著效果。喷施GA3各处理组与CK-2均未达到显著差异水平,由此可得,喷施GA3对低温胁迫下南美油藤幼苗叶片CAT活性值增加效果不显著。由于喷施GA3在处理24和48 h时叶片CAT活性变化均与CK-2呈显著差异(图7),因此可推断短时间低温胁迫下喷施GA3对幼苗叶片CAT活性的提高有一定的效果,但随着时间的延长,对CAT活性的增加效果不明显。
2. 4 生理生化指标间方差分析和相关性分析结果
利用Pearson相关系数法分析南美油藤幼苗不同生理生化指标间的相关性,结果如表8所示。南美油藤幼苗叶片REC与MDA含量呈正相关关系,即REC越高,细胞膜通透性越大,MDA含量越高,细胞膜功能越弱,南美油藤幼苗抗冷害能力越弱。SS含量与REC、MDA含量间均呈负相关关系,可能是SS在低温环境下增加了细胞的保水能力,降低了细胞质冰点,从而对幼苗起到保护作用。根据SS含量与其他指标间的相关系数可知,SS和SP含量与3个氧化酶活性指标均呈正相关关系;SP可提高细胞持水能力,保护原生质膜结构,提高细胞液浓度,从而降低冰点,提高植物抗冷害性,本研究中SP含量的变化与SS类似,对南美油藤幼苗抵抗冷害能力有积极的作用。SOD、POD和CAT共同作用能够清除细胞内自由基活性氧,其中SOD和POD能将有毒的自由基活性氧还原成H2O和O2,CAT能对膜脂和线粒体过氧化产生的活性氧有清除作用(任军等,2013)。本研究中上述3种保护性氧化酶间存在正相关关系,其活性的增加能提高南美油藤幼苗抵抗冷害的能力。从3种氧化酶活性与其他指标间的相关系数可知,POD活性与REC呈负相关关系,CAT活性与REC和MDA含量均呈负相关关系,由此可知,3种氧化酶活性的增加可适当降低南美油藤幼苗叶片REC和MDA含量,即3种氧化酶的活性增强可适当提高南美油藤幼苗叶片的持水能力,增强细胞膜的功能,提高南美油藤幼苗的抗冷害能力。
参考文献:
苍晶,王艳梅,王兴,于晶,王军虹,张达,张兆猛,樊立华. 2013. 根际浇灌ABA对冬小麦幼苗抗寒性的影响[J]. 东北农业大学学报,44(4): 36-42. [Cang J,Wang Y M,Wang X,Yu J,Wang J H,Zhang D,Zhang Z M,Fan L H. 2013. Effect of cold resistance on winter wheat seedlings with ABA pouring around root[J]. Journal of Nor-theast Agricultural University,44(4): 36-42.]
陈奇,袁金海,孙万仓,刘自刚,赵新旺,方彥,武军艳,李学才,曾秀存,米超,蒲媛媛,马骊,赵艳宁,方园,许耀照. 2017. 低温胁迫下白菜型冬油菜与春油菜叶片光合特性及内源激素变化比较[J]. 中国油料作物学报,39(1): 37-46. [Chen Q,Yuan J H,Sun W C,Liu Z G,Zhao X W,Fang Y,Wu J Y,Li X C,Zeng X C,Mi C,Pu Y Y,Ma L,Zhao Y N,Fang Y,Xu Y Z. 2017. Leaf photosynthetics and endogenous hormons of spring and winter rapa(Brassica rapa L.) under low temperature[J]. Chinese Journal of Oil Crop Sciences,39(1): 37-46.]
代勋,李忠光,龚明. 2012. 赤霉素、钙和甜菜碱对小桐子种子萌发及幼苗抗低温和干旱的影响[J]. 植物科学学报,30(2): 204-212. [Dai X,Li Z G,Gong M. 2012. Effect of gibberellin,calcium,and betaine on seed germination and resistance of Jatropha curcas L. seedling to low temperature and drought stress[J]. Plant Science Journal,30(2): 204-212.]
黄杏. 2012. 外源ABA提高甘蔗抗寒性的生理及分子机制研究[D]. 南宁:广西大学. [Huang X. 2012. Study on physio-logical and molecular mechanism of cold resistance enhanced by exogenous ABA application in sugarcane[D]. Nanning: Guangxi University.]
李学玲,庞海颖,牛东伟,任士福,李彦慧. 2017. 不同外源激素对李花抗寒性及相关生理指标的影响[J]. 北方园艺,(11): 17-22. [Li X L,Pang H Y,Niu D W,Ren S F,Li Y H. 2017. Effect of different exogenous hormones on cold-resistant ability and related physiological indicators of Plum flower[J]. Northern Horticulture,(11): 17-22.]
李馨园,杨晔,张丽芳,左师宇,李丽杰,焦健,李晶. 2017. 外源ABA 对低温胁迫下玉米幼苗内源激素含量及Asr1基因表达的调节[J]. 作物学报,43(1): 141-148. [Li X Y,Yang Y,Zhang L F,Zuo S Y,Li L J,Jiao J,Li J. 2017. Regulation on contents of endogenous hormones and Asr1 gene expression of maize seedling by exogenous ABA under low-temperature stress[J]. Acta Agronomica Sinica,43(1): 141-148.]
刘果,陈鸿鹏,彭彦,谢耀坚,陈少雄. 2018. 印加果种子生长发育过程中脂肪酸组成的变化分析[J]. 中国油脂,43(8): 57-62. [Liu G,Chen H P,Peng Y,Xie Y J,Chen S X. 2018. Variations in fatty acid compasition of Sacha inchi seeds during growth and development[J]. China Oils and Fats,43(8): 57-62.]
刘果,陈鸿鹏,吴志华,彭彦,谢耀坚. 2019. 南美油藤种子发育过程的代谢组学和转录组学联合分析[J]. 林业科学,55(5): 169-179. [Liu G,Chen H P,Wu Z H,Peng Y,Xie Y J. 2019. Analyses of seed development of Plukenetia volubilis by joint metabolomics and transcriptomics approaches[J]. Scientia Silvae Sinicae,55(5): 169-179.]
罗银玲,苏志龙,崔现亮,李孙洋,兰芹英. 2014. 外源ABA提高星油藤幼苗抗冷害能力的探讨[J]. 云南大学学报(自然科学版),36(6): 936-941. [Luo Y L,Su Z L,Cui X L,Li S Y,Lan Q Y. 2014. An investigation of exogenous abscisic acid on improving chilling tolerance of the seedlings of sacha inchi(Plukenetia volubilis)[J]. Journal of Yunnan University(Natural Sciences Edition),36(6): 936-941.]
羅正荣. 1989. 植物激素与抗寒力的关系[J]. 植物生理学通讯,(3): 1-5. [Luo Z R. 1989. Relationship between plant hormones and cold resistance[J]. Plant Physiology Journal,(3): 1-5.]
聂庆娟,孟朝,梁海永,孟庆瑞,李彦慧. 2007. 低温胁迫对4种常绿阔叶植物膜脂过氧化及保护酶活性的影响[J]. 植物研究,27(5): 578-581. [Nie Q J,Meng C,Liang H Y,Meng Q R,Li Y H. 2007. Effect on low temperature stress on lipid peroxidation and activity of cell defense enzymes of four evergreen broadleaved tree species[J]. Bulletin of Botanical Research,27(5): 578-581.]
任军,黄志霖,曾立雄,施征. 2013. 低温胁迫下植物生理反应机理研究进展[J]. 世界林业研究,26(6): 15-20. [Ren J,Huang Z L,Zeng L X,Shi Z. 2013. A review of physio-logical reaction mechanism of plants exposed to low temperature stress[J]. World Forestry Research,26(6): 15-20.]
苏鹏. 2010. 不同草坪草的温度胁迫抗性差异及其机理研究[D]. 长沙:湖南农业大学. [Su P. 2010. Studies on the differences of temperature stressed tolerance and mechanism in diverse turfgrass species[D]. Changsha: Hunan Agricultural University.]
王翠花,刘沙,张瑞富,毕文波,郭志富. 2014. 植物抗寒分子生物学研究概况及展望[J]. 辽宁农业科学,(1): 45-48. [Wang C H,Liu S,Zhang R F,Bi W B,Guo Z F. 2014. Overview and prospect of plant cold resistance molecular biology research[J]. Liaoning Agricultural Sciences,(1): 45-48.]
王连荣,薛拥志,常美花,王莉. 2016. 外源激素对杏扁抗寒生理指标的影响[J]. 核农学报,30(2): 396-403. [Wang L R,Xue Y Z,Chang M H,Wang L. 2016. Effects of exo-genous hormones on cold resistant physiological index of Kernel Apricot[J]. Journal of Nuclear Agricultural Scien-ces,30(2): 396-403.]
魏鑫,倪虹,张会慧,王晶英. 2016. 外源脱落酸和油菜素内酯对干旱胁迫下大豆幼苗抗旱性的影响[J]. 中国油料作物学报,38(5): 605-610. [Wei X,Ni H,Zhang H H,Wang J Y. 2016. Effects of exogenous abscisic acid and brassinolide on drought resistance of soybean seedlings[J]. Chinese Journal of Oil Crop Sciences,38(5): 605-610.]
闫绍鹏. 2004. 欧美山杨杂种无性系低温胁迫下几种生理指标的遗传变异[D]. 哈尔滨:东北林业大学. [Yan S P. 2004. Genetic variation among the hybrid clones of P. tremula×P. tremuloides in several physiological characteri-stics under low temperature stress[D]. Harbin: Northeast Forestry University.]
Achard P,Renou J P,Berthomé R,Harberd N P,Genschik P. 2008. Plant DELLAs restrain growth and promote survi-val of adversity by reducing the levels of reactive oxygen species[J]. Current Biology,18(9): 656-660.
Alicia A N,Isabella E J. 2016. Sacha Inchi(Plukenetia volubilis Linneo): A wasted ancestral experience? Clinical evidence related to its consumption[J]. Revista Chilena de Nutrición,43(2): 167-171.
Chandrasekaran U,Liu A Z. 2015. Stage-specific metabolization of triacylglycerols during seed germination of Sacha Inchi(Plukenetia volubilis L.)[J]. Journal of the Science of Food and Agriculture,95(8): 1764-1766.
de Souza A H P,Gohara A K,Rodrigues A C,de Souza N E,Visentainer J V and Matsushita M. 2013. Sacha inchi as potential source of essential fatty acids and tocopherols: Multivariate study of nut and shell[J]. Acta Scientiarum,35(4): 757-763.
Hu X D,Pan B Z,Fu Q T,Niu L J,Chen M S,Xu Z F. 2018. De novo transcriptome assembly of the eight major organs of Sacha Inchi(Plukenetia volubilis) and the identification of genes involved in α-linolenic acid metabolism[J]. BMC Genomics. doi:10.1186/s12864-018-4774-y.
Jones R L. 1973. Gibberellins: Their physiological role[J]. Annual Review of Plant Physiology,24: 571-598.
Kumar B,Smita K,Cumbal L,Debut A. 2017. Sacha inchi(Plukenetia volubilis L.) shell biomass for synthesis of silver nanocatalyst[J]. Journal of Saudi Chemical Society. doi:10.1016/j.jscs.2014.03.005.
Rakei A,Maali-Amiri R,Zeinali H,Ranjbar M. 2016. DNA methylation and physio-biochemical analysis of chickpea in response to cold stress[J]. Protoplasma,253: 61-76.
Richards D E,King K E,Ait-Ali T,Harberd N P. 2001. How gibberellin regulates plant growth and development: A molecular genetic analysis of gibberellin signaling[J]. Annual Review of Plant Biology. doi:10.1146/annurev.arplant.52.1.67.
Uzilday B,Turkan I,Sekmen A H,Ozgur R,Karakaya H C,2012. Comparison of ROS formation and antioxidant enzymes in Cleome gynandra(C4) and Cleome spinosa(C3) under drought stress[J]. Plant Science,182: 59-70.
Wang F,Liang D Y,Pei X N,Zhang Q H,Zhang P,Zhang J Q,Lu Z M,Yang Y C,Liu G F,Zhao X Y. 2019. Study on the physiological indices of Pinus sibirica and Pinus koraiensis seedlings under cold stress[J]. Journal of Fo-restry Research. doi:10.1007/s11676-018-0833-0.
Zhu Z,Ding Y,Zhao J H,Nie Y,Zhang Y,Liu C,Tang X M. 2016. Effects of postharvest gibberellic acid treatment on chilling tolerance in cold-stored tomato(Solanum lycopersicum L.) fruit[J]. Food and Bioprocess Technology. doi:10.1007/s11947-016-1712-3.
(責任编辑 邓慧灵)
猜你喜欢
安徽农学通报(2017年2期)2017-04-25
江苏农业科学(2017年1期)2017-02-27
江苏农业科学(2016年6期)2016-07-25
江苏农业科学(2015年8期)2015-09-10
江苏农业科学(2015年8期)2015-09-10
安徽农学通报(2014年24期)2015-01-08
湖北农业科学(2014年20期)2014-12-12
热带农业科学(2014年8期)2014-09-23
安徽农学通报(2014年12期)2014-07-28
