
贵州小麦品种(系)粒重相关基因TaCwi-A1、TaSus2-2B和TaGW2-6A等位变异类型鉴定
2020-12-11 09:10杨雪敏李鲁华任明见徐如宏
南方农业学报 2020年9期
杨雪敏 李鲁华 任明见 徐如宏

摘要:【目的】鑒定贵州小麦品种(系)中粒重相关基因TaCwi-A1、TaSus2-2B和TaGW2-6A的等位变异类型,筛选含高粒重基因型的小麦品种(系),为贵州小麦粒重的遗传改良和高粒重品种选育提供参考。【方法】以252份小麦品种(系)为材料,分别利用TaCwi-A1、TaSus2-2B和TaGW2-6A基因的分子标记(CWI22/CWI21、TaSus2-1/TaSus2-2和Hap-6A-P1/Hap-6A-P2)引物进行PCR扩增,利用毛细管电泳检测扩增产物,鉴定分析这3个基因的等位变异类型及分布频率,并结合籽粒性状测定结果,筛选含高粒重基因变异类型的小麦种质。【结果】252份小麦种质材料的粒重平均值为39.87 g,其中,有56份材料属于大粒种质(>45.00 g),仅有13份检测到等位变异;173份材料属于中粒种质(30.00~45.00 g),有24份检测到等位变异;23份材料属于小粒种质(<30.00 g),均未检测到等位变异。252份小麦材料中,含有TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异的材料37份,占供试材料总数的14.7%,其中TaCwi-A1基因等位变异类型材料12份(包括TaCwi-A1a变异类型8份,TaCwi-A1b变异类型4份),占供试材料总数的4.8%;TaSus2-2B基因等位变异类型材料16份(包括TaSus2-2BH变异类型材料2份,TaSus2-2BL变异类型材料14份),占供试材料总数的6.4%;TaGW2-6A基因等位变异类型材料10份(包括Hap-6A-A变异类型4份,Hap-6A-G变异类型6份),占供试材料总数的4.0%;等位变异组合类型仅有1份材料(惠光2-2-2),为TaCwi-A1b/HAP-6A-A,占供试材料总数的0.4%。平均粒重最高的变异类型为TaCwi-A1b,其次是HAP-6A-G变异类型。对于平均粒重,TaCwi-A1b变异类型显著高于TaCwi-A1a变异类型(P<0.05,下同),Hap-6A-G变异类型显著高于Hap-6A-A变异类型,TaSus2-2BH变异类型也高于TaSus2-2BL变异类型,但差异不显著(P>0.05)。【结论】从贵州小麦品种(系)检测到的粒重基因等位变异整体较少,表明贵州小麦种质遗传多样性较低,高粒重品种的选育工作开展不够,今后应重视小麦粒重基因等位变异综合效应研究。鉴定出含有TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异类型、粒重>45.00 g的品种(系)13份,可应用于贵州小麦粒重的遗传改良和高粒重品种选育。
关键词: 小麦;粒重;基因;等位变异;分子标记;毛细管电泳;鉴定
中图分类号: S512.103.53 文献标志码: A 文章编号:2095-1191(2020)09-2071-11
Identification and analysis of allele variation types of kernel weight related genes TaCwi-A1,TaSus2-2B and TaGW2-6A of Guizhou wheat varieties(lines)
YANG Xue-min, LI Lu-hua, REN Ming-jian, XU Ru-hong*
(College of Agriculture, Guizhou University/Guizhou Branch of National Wheat Improvement Center,
Guiyang 550025,China)
Abstract:【Objective】The allele variation types of kernel weight related genes TaCwi-A1,TaSus2-2B and TaGW2-6A in Guizhou wheat varieties(lines) were identified,and wheat varieties(lines) containing high kernel weight genotypes were screened to provide reference for genetic improvement of Guizhou wheat kernel weight and breeding of high kernel weight varieties. 【Method】With 252 wheat varieties(or lines) as the materials,using molecular marker of genes TaCwi-A1, TaSus2-2B and TaGW2-6A(CWI22/CWI21, TaSus2-1/TaSus2-2 and Hap-6A-P1/Hap-6A-P2) primers for PCR amplification,amplification products were detected using capillary electrophoresis,allele variation types and distribution of the three genes were identified, combined with the measuring results of grain traits, wheat germplasm with high kernel weight mutation types were screened. 【Result】The average grain weight of 252 wheat germplasm materials was 39.87 g. Among them,56 materials belonged to large kernel germplasms(>45.00 g),and only 13 of them detected allele variation.Among the 173 medium kernel materials(30.00-45.00 g),allele variation was detected in 24. The rest 23 materials belonged to small kernel germplasm(<30.00 g),and no allele variation was detected.Among the 252 wheat materials,37 contained allelic variation of TaCwi-A1, TaSus2-2B and TaGW2-6Agenes,accounting for 14.7% of the total tested mate-rials. Among them,12 contained allele variation of TacWI-A1 gene(including 8 of TaCwi-A1a and 4 of TaCwi-A1b),accou-nting for 4.8% of the total tested materials. There were 16 allele materials of TaSus2-2B gene(including 2 allele materials of TaSus2-2BH and 14 allele materials of TaSus2-2BL),accounting for 6.4% of the total number of tested materials. There were 10 allele variation types of TAGW2-6A(including 4 HAP-6A-A variation types and 6 HAP-6A-G variation types),accou-nting for 4.0% of the total number of materials tested.There was only one allele variation combination type of material(Huigang 2-2-2),which was Tacwi-A1b/HAP-6A-A,accounting for 0.4% of the total number of materials tested. The varia-tion type with the highest mean kernel weight was TACwi-A1b,the second type was HAP-6A-G variation. For average grain weight,the variation type of Tacwi-A1b was significantly higher than that of Tacwi-A1a(P<0.05,the same as below),the variation type of HAP-6A-G was significantly higher than that of HAP-6A-A,and the variation type of TaSus2-2BH was also higher than that of TaSus2-2BL,but the difference was not significant(P>0.05). 【Conclusion】The allele variation diversity of grain weight genes detected from Guizhou wheat varieties(lines) is relatively low,indicating that the genetic diversity of Guizhou wheat germplasm is low,and the breeding of high-kernel-weight varieties is not enough. In the future,more attention should be paid to the comprehensive effects of the allele variation of wheat weight. Thirteen varie-ties(lines) containing allele types of TaCwi-A1,TaSus2-2B and TaGW2-6A genes with grain weight of >45.00 g were identified,which can be used for genetic improvement of Guizhou wheat grain weight and breeding of high-kernel-weight varie-ties.
Key words: wheat; kernel weight; gene; allele variation; molecular markers; capillary electrophoresis; identification
Foundation item:National Key Research and Development Plan “Seven Crops Breeding” Key Special Project(2017YFD0100900);Introducing Talents Research Project of Guizhou University(GDRJHZ〔2017〕49)
0 引言
【研究意义】小麦(Triticum aestivum L.)是重要的粮食作物,全球约40%的人口以其为主食(http://faost at3.fao.org/home)。小麦高效生产才能实现全球粮食和营养安全(Hunter et al.,2017)。粒重是小麦产量的主要构成因素,当小麦粒重增加1 g时,每公顷小麦产量可增加140~160 kg,因此,粒重是小麦产量的重要影响因素,培育高粒重品种是小麦育种的重要目标(Tian et al.,2006;Dong et al.,2012)。贵州生态环境复杂,处于气候潮湿多雨的西南麦区,小麦粒重在不同年度间差异明显,且粒重是受多个微效基因控制的数量性状,仅靠表型选择聚合粒重优异等位基因较困难(刘永伟等,2017a)。借助分子标记技术,对影响粒重的相关基因进行分子标记辅助选择,可极大提高贵州小麦产量育种的效率。因此,利用粒重相关基因的功能标记对小麦性状改良及高产育种具有重要意义(司文洁等,2019)。【前人研究进展】关于小麦粒重相关基因的研究已被广泛报道,尤其以粒重相关基因TaCwi-A1、TaSus2-2B和TaGW2-6A的研究较多。韩利明等(2011)利用小麦粒重基因TaGW2-6A位点的CAPS分子标记检测来自21个国家的745份小麦种质材料,结果发现TaGW2-6A位点的CAPS分子标记能很好区分其等位变异类型Hap-6A-A和Hap-6A-G,可作为粒重辅助选择的有效位点;Jiang等(2011)克隆得到小麦TaSus2基因,并证明该基因与小麦产量性状相关联,其在胚乳发育过程中表达量较高;Su等(2011)通过同源克隆方法从小麦中获得水稻OsGW2基因的同源基因TaGW2,研究发现该基因对小麦粒重起负调控作用;Ma等(2012)也采用同源克隆方法获得小麦TaCwi-A1基因的全长cDNA,并对其进行QTL分析,證明其可解释粒重4.8%的表型差异;相吉山等(2014)、刘永伟等(2017b)研究发现,含有TaCwi-A1a基因的小麦粒重显著高于含有TaCwi-A1b基因的小麦粒重;刘永伟等(2017a)研究发现,TaGW2-6A、TaCwi-A1和TaSus2-2B基因与产量相关,对小麦粒宽、粒重等重要产量性状均发挥调控作用,直接影响小麦的产量;时佳等(2018)、仝靖洋等(2018)利用TaGW2-6A、TaCwi-A1和TaSus2-2B基因的分子标记研究其与小麦粒重的相关性,结果发现这些标记均能有效预测小麦粒重大小,可用于小麦粒重的分子辅助选择研究。【本研究切入点】目前有关利用分子标记鉴定贵州小麦TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异类型的研究鲜见报道。【拟解决的关键问题】将分子标记和毛细管电泳检测技术相结合,对252份小麦品种(系)中TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异类型进行鉴定,筛选含高粒重基因型的小麦种质,为小麦高粒重品种选育和分子辅助选择研究提供参考。
1 材料与方法
1. 1 试验材料
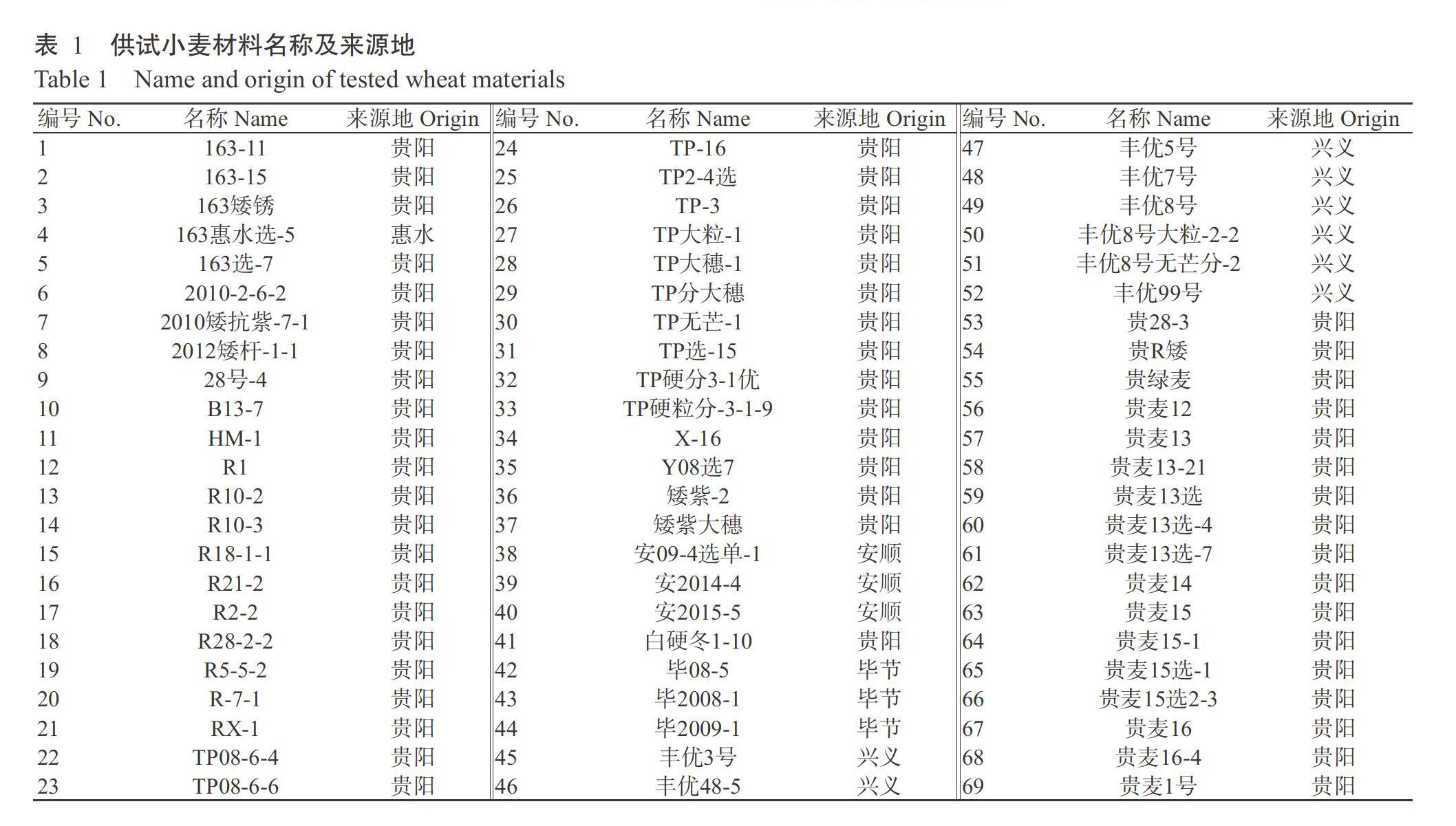
供试材料为252个小麦品种(系),分别来自贵州贵阳、兴义、毕节、安顺和惠水等地,其中部分材料由国家小麦改良中心贵州分中心选育,名称和来源见表1。主要试剂:DNF-910 dsDNA Kit/s试剂盒购自成都百乐科技有限公司;2×Taq PCR Master Mix反应试剂购自生工生物工程(上海)股份有限公司;DL2000 DNA Marker购自贵州弥勒天根生物科技有限公司;905-33-DNA-0~500 bp Mthds试剂盒购于研诺逻辑集成电路设计有限公司;50×TAE溶液购自北京索莱宝科技有限公司。主要设备仪器:移液器(Eppendorf,德国)、Genova Nano分光光度计(Jenway,英国)、T100TM_Thermal Cycler PCR仪(Bio-Rad,美国)、Fragment Analyzer毛细管电泳系统(AATI,美国)、高速离心机(Thermo,美国)、水平电泳槽(DYCP-32B,北京六一仪器厂)、电泳仪(DYY-6C,北京六一生物技术有限公司)、电热恒温水浴锅(DK-98-Ⅱ,天津泰斯特仪器有限公司)、高速冷冻离心机(Thermo,美国)和摇床(GS-20,杭州米欧仪器有限公司)。
1. 2 试验方法
1. 2. 1 小麦籽粒性状测定 于2017─2018年将供试小麦材料种植于国家小麦改良中心贵州分中心科研基地。成熟期,每份材料随机选取500粒左右的种子,用SC-G型多功能种子分析仪测量粒重、粒长和粒宽,每份材料的每一项指标均重复测量3次,计算平均值。
1. 2. 2 基因组DNA提取 每份材料随机取10粒种子进行发芽,采用植物基因组DNA提取试剂盒提取其DNA,以分光光度计检测其浓度,并稀释至50 ng/μL备用。
1. 2. 3 分子标记检测 利用前人已开发的分子标记检测TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位变异,所用分子标记引物的相关信息如表2所示,由生工生物工程(上海)股份有限公司合成。以上述提取的DNA为模板进行PCR扩增。反应体系20.0 μL:10×PCR Buffer 2.0 μL,2.5 mmol/L dNTPs 0.4 μL,25 mmol/L MgCI2 1.6 μL,10 μmol/L正、反向引物各1.0 μL,2.5 U/μL Taq DNA聚合酶0.5 μL,50 ng/μL DNA模板2.0 μL,用ddH2O补足至20.0 μL。将反应体系置于T100 PCR仪上进行PCR扩增。扩增程序:94 ℃预变性3 min;94 ℃ 30 s,退火温度和时间见表2,72 ℃ 1 min,进行30个循环;72 ℃延伸10 min。
Hap-6A-P1和Hap-6A-P2为CAPS分子标记,参照Su等(2011)的方法检测TaGW2-6A基因的这2个CAPS分子标记:用Hap-6A-P1引物进行第一轮PCR扩增,然后将第一轮扩增的PCR产物适当稀释后作为模板,用Hap-6A-P2引物进行第二轮PCR扩增。第二轮PCR扩增完毕后,用限制性内切酶Taq I酶切第二轮PCR扩增产物,PCR产物和酶切片段分别用毛细管电泳进行分离。
1. 2. 4 毛细管电泳检测 上述PCR扩增产物使用Fragment AnalyzerTM毛细管电泳系统进行检测及分离。取2.0 μL PCR扩增产物,加入22.0 μL Buffer TE。检测程序:预运行电压和时间是6 kV和30 s;Marker进样电压和时间为5 kV和10 s;样品进样电压和时间分别是5 kV和10 s;分离时电压和时间是6 kV和45 min。
1. 3 统计分析
使用SPSS和PROSize 3.0进行数据统计分析。粒重、粒长、粒宽等试验数据使用SC-G型多功能种子分析仪进行独立样本检验。扩增结果使用PRO-Size 3.0进行统计分析。
2 结果与分析
2. 1 小麦籽粒表型性状测定结果
252份小麦材料的粒重平均为39.87 g,粒长平均为6.89 mm,粒宽平均为3.07 mm,其中以Y08选7的粒重最大(57.93 g),以中2012-14的粒重最小(21.55 g)(表3)。根据李立会和李秀全(2006)的小麦粒重划分标准,有56份材料属于大粒种质(>45.00 g),173份材料属于中粒种质(30.00~45.00 g),23份材料属于小粒种质(<30.00 g)。从变异系数来看,以粒重的变异系数较大,为16.8%,表明252份小麦品种的粒重遗传变异较大。总体来说,252份小麦材料中以低粒重品种居多,说明对贵州小麦品种(系)进行粒重鉴定及选育改良非常必要。
2. 2 TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异类型检测结果
对252份小麦材料TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位变异类型检测结果如表3所示。56份大粒种质中仅有13份(ZY96-3、贵农10-6、贵麦15、贵紫2号、贵农10-6-16-2、贵农10-6-16-6、贵农10-6-16-7、贵农15、贵农15选-2-4、TP无芒-1、丰优99号、贵麦15-1和惠光2-2-2)检测到等位变异类型,这些种质可用于贵州小麦粒重的遗传改良和高粒重品种选育,173份中粒种质中有24份检测到等位变异类型,23份小粒种质中均未检测到等位变异类型。
2. 2. 1 TaCwi-A1等位变异类型检测结果 根据Ma等(2012)的研究报道,TaCwi-A1基因存在2种等位变异类型,其中TaCwi-A1a变异类型用CWI22标记引物能扩增出402 bp的目的片段,TaCwi-A1b变异类型用CWI21标记引物能扩增出404 bp的目的片段。对CWI22和CWI21标记引物的扩增产物进行毛细管电泳检测,结果发现,252份小麦材料中有8份材料(丰优99、贵麦15、贵农10-6、贵农12-7、贵农33、贵优麦、兴义98-2-4和ZY96-3)能扩增出402 bp的目的片段(表3和图1),说明这8份材料为TaCwi-A1基因的TaCwi-A1a變异类型,占供试材料总数的3.2%;有4份材料(TP无芒-1、贵农10-6-16-6、贵紫2号和惠光2-2-2)能扩增出404 bp的目的片段(表3和图2),说明这4份材料为TaCwi-A1基因的TaCwi-A1b变异类型,占供试材料的1.6%。其他材料未检测到到这2个目的片段。
2. 2. 2 TaSus2-2B基因等位变异类型检测结果 根据Jiang等(2011)的研究报道,TaSus2-2B基因存在2种等位变异类型,其中,TaSus2-2BH变异类型用TaSus2-2BH标记引物能扩增出423 bp的目的片段,TaSus2-2BL变异类型用TaSus2-2BL标记引物能扩增出381 bp的目的片段。对TaSus2-1和TaSus2-2标记引物的PCR扩增产物进行毛细管电泳检测,252份小麦材料中有2份材料(贵农10-19和贵紫4号)能扩增出423 bp的目的片段(表3和图3),说明这2份材料为TaSus2-2B基因的TaSus2-2BH变异类型,占供试材料总数的0.8%;有14份材料(163-15、TP-16、TP-3、贵麦15、贵农08-9选10、贵农10-3-16-9、贵农10-4-16-20、贵农12、贵农15、贵农15选-2-4、惠TP分-1、节四燕、张08-6特优-2和张10选-4-1)能扩增出381 bp的目的片段(表3和图4),说明这14份材料为TaSus2-2B基因的TaSus2-2BL变异类型,占供试材料总数的5.6%。其他材料未检测到这2个目的片段。
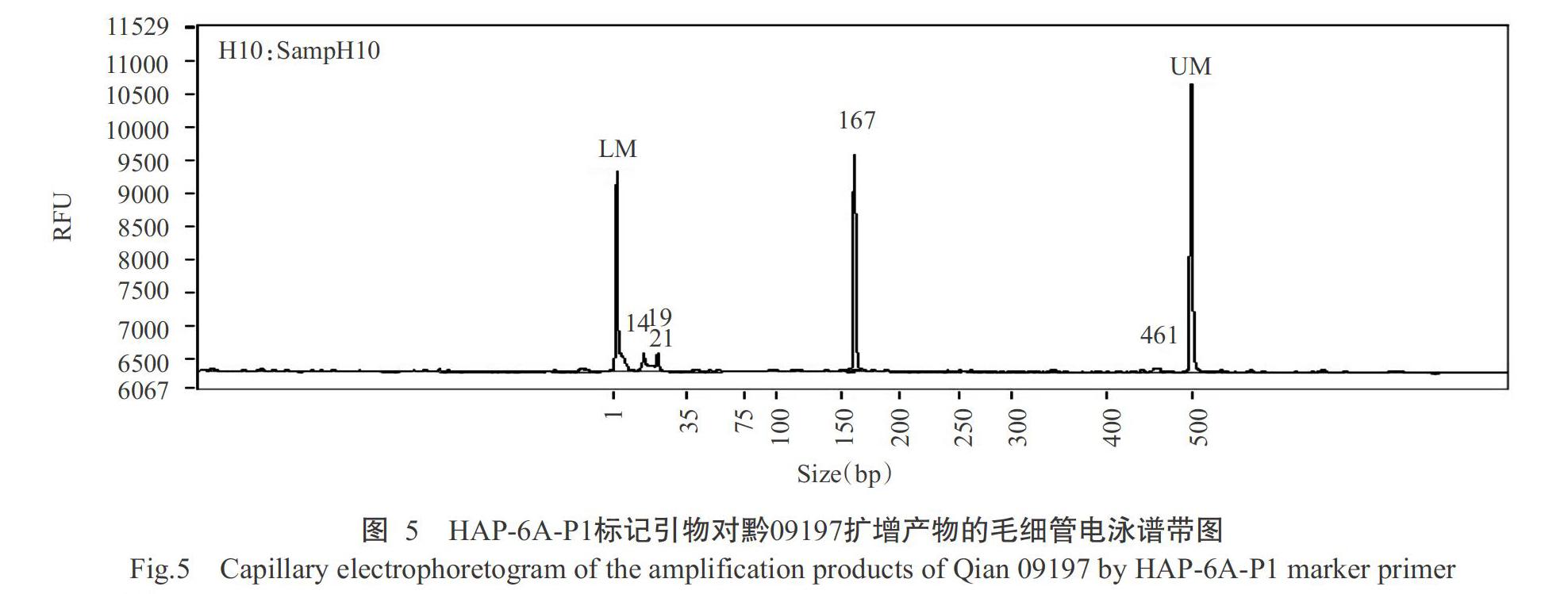
2. 2. 3 TaGW2-6A基因等位变异类型检测结果 根据Su等(2011)的研究报道,TaGW2-6A基因利用相应的CAPS分子标记能检测到Hap-6A-A和Hap-6A-G 2种等位变异类型。其中Hap-6A-A变异类型能检测到167 bp的目的片段,Hap-6A-G变异类型能检测到218 bp的目的片段。对Hap-6A-P1和Hap-6A-P2标记引物的PCR扩增产物进行毛细管电泳检测,结果发现,252份材料中有4份材料(贵农10-18、贵农30-16、惠光2-2-2和黔09197)能检测到167 bp的目的片段(表3和图5),说明这4份材料为TaGW2-6A基因的Hap-6A-A变异类型,占供试材料总数的1.6%;有6份材料(贵农10-6-16-9、贵农10-6-16-2、贵农10-6-16-7、贵农31、贵农优麦2号和惠紫1号)能检测到218 bp的目的片段(表3和图6),说明这6份材料为TaGW2-6A基因的Hap-6A-G变异类型,占供试材料总数的2.4%。其他材料未检测到这2个目的片段。
2. 3 等位变异类型统计及籽粒表型差异显著性分析结果
TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位变异类型统计结果如表4所示。252份小麦材料中,含有TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异的材料有37份,占供试材料总数的14.7%,其中TaSus2-2B基因等位变异类型材料16份,占供试材料总数的6.4%,TaCwi-A1基因等位变异类型材料有12份,占供试材料总数的4.8%,TaGW2-6A基因等位变异类型材料有10份,占供试材料总数的4.0%;等位变异组合类型仅有1份材料(惠光2-2-2),为TaCwi-A1b/HAP-6A-A,占供试材料总数的0.4%;有43份粒重高于45.00 g的材料均未检测到TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异。平均粒重最重的变异类型为TaCwi-A1b,为47.60 g,其次是HAP-6A-G变异类型,平均粒重为44.04 g。
由表4还可知,TaCwi-A1b变异类型材料的平均粒重(47.60 g)显著高于TaCwi-A1a(41.95 g)变异类型(P<0.05,下同),Hap-6A-G變异类型材料的平均粒重(44.04 g)显著高于Hap-6A-A变异类型(40.77 g),说明TaCwi-A1b和Hap-6A-G变异类型均能明显增加小麦的粒重。TaSus2-2BH变异类型材料的平均粒重(43.47 g)也高于TaSus2-2BL变异类型(41.75 g),但差异不显著(P>0.05,下同)。在TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位变异类型中,仅TaSus2-2BH变异类型材料的平均粒长(7.21 mm)显著高于TaSus2-2BL变异类型材料(7.05 mm),其他变异类型材料的平均粒长和平均粒宽差异均不显著。
3 讨论
本研究对贵州252份种质材料中粒重相关基因(TaCwi-A1、TaSus2-2B和TaGW2-6A)等位变异的类型及分布频率进行研究,结果发现14.7%的供试材料含有TaCwi-A1a、TaCwi-A1b、TaSus2-2BH、TaSus2-2BL、HAP-6A-A和HAP-6A-G 6种变异类型,其中以TaSus2-2BL变异类型材料最多,占供试材料总数的5.6%,以TaSus2-2BH变异类型材料最少,占供试材料总数的0.8%,且从所有材料中只检测到1份材料为等位变异组合类型TaCwi-A1b/HAP-6A-A,仅占供试材料总数的0.4%。与前人研究结果相比,本研究中各基因等位变异类型所占比例均较低。如刘永伟等(2017b)研究发现,黄淮麦区小麦中TaCwi-A1a变异类型所占比列为65.03%;李娜等(2018)研究显示,在64个青海小麦品种中,62.5%的材料为HAP-6A-A变异类型;时佳等(2018)研究发现,219份新疆小麦种质材料中,40.18%的材料为TaSus2-B2A变异类型;仝靖洋等(2018)从262份供试材料中检测出8种等位变异组合类型,远多于本研究检测出的等位变异组合类型。存在较大差异的原因除了供试材料不同外,很可能是本研究中的供试材料主要是地方品种,高代相近品系较多,整体遗传多样性较低,导致检测到的粒重基因等位变异较少。这反映出贵州小麦高粒重品种的选育工作开展不够,今后应重视小麦粒重基因等位变异综合效应研究。
本研究还发现,TaSus2-2BH变异类型材料的平均粒重高于TaSus2-2BL变异类型材料,与Jiang等(2011)、Ma等(2012)的研究结果一致;TaCwi-A1b变异类型材料的平均粒重显著高于TaCwi-A1a变异类型材料,与Jiang等(2011)、Ma等(2012)的研究结果存在差异;Hap-6A-G变异类型材料的平均粒重高于Hap-6A-A变异类型材料,与李娜等(2018)的研究结果相同,而与Su等(2011)的研究结果不同;在TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位变异类型中,仅TaSus2-2BH变异类型材料的平均粒长显著高于TaSus2-2BL变异类型材料,其他变异类型材料的平均粒长和平均粒宽差异均不显著,与仝靖洋等(2018)的研究结果相似,而与刘永伟等(2017b)的结果不同。上述与前人研究结果不一致的主要原因是供试材料中检测到的各等位变异类型材料份数较少,且测定的粒重仅为1年数据,不同等位变异类型对不同材料粒重的贡献率仅作为参考,今后应扩大样本量,进一步开展不同变异类型的综合效应研究,用于小麦高粒重品种分子辅助选择育种。此外,本研究发现43份粒重高于45.00 g的材料均未检测到TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异,针对这些材料可开展多年多点试验鉴定是否含有其他粒重相关的新基因,将这些新基因应用于小麦高粒重品种遗传改良。
据报道,贵州小麦育种在“十一五”期间一直徘徊不前,如果还是在现有的遗传背景下继续进行品种选育,则很难有重大突破(英敏和余虎,2012),应通过拓宽高粒重的亲本遗传背景来提高小麦品种的产量潜力(何昀昆等,2008)。大量研究证实,TaCwi-A1a、TaSus2-B2H等均为优异等位变异类型,其对粒重有显著增效作用,今后可加大聚合多个优异等位变异组合类型的种质资源收集、鉴定和筛选研究,提高其在贵州小麦种质资源中的比例,开展小麦高粒重品种的遗传改良,从而有效提高贵州小麦产量整体水平。
4 结论
从贵州小麦品种(系)检测到的粒重基因等位变异整体较少,表明贵州小麦种质遗传多样性较低,高粒重品种的选育工作开展不够,今后应重视小麦粒重基因等位变异综合效应研究。鉴定出含有TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位变异类型、粒重>45.00 g的品种(系)13份,可应用于贵州小麦粒重的遗传改良和高粒重品种选育。
参考文献:
韩利明,杨芳萍,夏先春,阎俊,张勇,曲延英,王忠伟,何中虎. 2011. 株高、粒重及抗病相关基因在不同国家小麦品种中的分布[J]. 麦类作物学报,31(5):824-831. [Han L M,Yang F P,Xia X C,Yan J,Zhang Y,Qu Y Y,Wang Z W,He Z H. 2011. Distribution of plant height,grain weight and disease-resistance related genes in wheat varie-ties in different countries[J]. Journal of Triticeae Crops,31(5):824-831.]
何昀昆,田晓琴,何庆才. 2008. 贵州省小麦育种研究与分析[J]. 种子,(10):95-97. [He Y K,Tian X Q,He Q C. 2008. Research and analysis of wheat breedingin Guizhou[J]. Seed,(10):95-97.]
李立会,李秀全. 2006. 小麦种质资源描述规范和数据标准[M]. 北京:中国农业出版社. [Li L H,Li X Q. 2006. Descriptors and data standard for wheat[M]. Beijing:China Agricultural Press.]
李娜,魏乐,曹东,陈文杰,张波,张怀刚,刘宝龙. 2018. 来源于人工合成小麦的TaGW2-6A等位变异类型的鉴定及分析[J]. 分子植物育种,16(2):518-525. [Li N,Wei L,Cao D,Chen W J,Zhang B,Zhang H G,Liu B L. 2018. Identification and analysis of TaGW2-6A allelic variation from synthetic wheat[J]. Molecular Plant Breeding,16(2):518-525.]
刘永伟,周硕,王雪征,孙果忠,董福双,柴建芳,李春杰,赵和,王海波. 2017a. 小麦粒重相关分子标记Xgwm46等位变异的鉴定与评价[J]. 麦类作物学报,37(1):7-15. [Liu Y W,Zhou S,Wang X Z ,Sun G Z,Dong F S,Chai J F,Li C J,Zhao H,Wang H B. 2017a. Identification and evaluation of molecular marker Xgwm46 allelic variations associated with kernel weight in wheat[J]. Journal of Triticeae Crops,37(1):7-15.]
刘永伟,周硕,王雪征,孙果忠,朱金永,韩秋芬,李春杰,赵和,王海波. 2017b. 粒重基因TaCwi-A1等位变异在黄淮麦区小麦品种(系)中的分布及功能分析[J]. 华北农学报,32(2):131-137. [Liu Y W,Zhou S,Wang X Z,Sun G Z,Zhu J Y,Han Q F,Li C J,Zhao H,Wang H B. 2017b. Functional analysis and distribution of allelic varia-tions of TaCwi-A1 gene related to kernel weight in Ye-llow and Huai river valleys facultative wheat zone[J]. Acta Agriculturae Boreali-Sinica,32(2):131-137.]
时佳,白璐,任毅,穆培源,梁晓东,玛依拉,耿洪伟. 2018. 新疆小麦TaGW2-6A、TaCwi-A1、TaSus2-2B等位变异对粒重的影响及应用[J]. 分子植物育种,16(3):848-858. [Shi J,Bai L,Ren Y,Mu P Y,Liang X D,Ma Y L,Di H W. 2018. Effects and application of allelic variation of TaGW2-6A,TaCwi-A1 and TaSus2-2B on grain weight of Xinjiang wheat[J]. Molecular Plant Breeding,16(3):848-858.]
司文洁,吴林楠,郭利建,周梦蝶,刘香利,马猛,赵惠贤. 2019. 小麦粒重相关基因TaCYP78A5功能标记开发及验证[J]. 作物学报,45(12):1905-1911. [Si W J,Wu L N,Guo L J,Zhou M D,Liu X L,Ma M,Zhao H X. 2019. Development and validation of the functional marker of grain weight-related gene TaCYP78A5 in wheat(Triticum aestivum L.)[J]. Acta Agronomica Sinica,45(12):1905-1911.]
仝靖洋,李少鵬,刘胜杰,张琳雪,梁园园,张哲,聂小军,李学军,王中华,高欣. 2018. 小麦粒重基因等位变异的高通量分子检测及组合分析[J]. 麦类作物学报,38(11):1300-1308. [Tong J Y,Li S P,Liu S J,Zhang L X,Liang Y Y,Zhang Z,Nie X J,Li X J,Wang Z H,Gao X. 2018.High-throughput molecular detection and analysis of allelic variation combinations related to grain weight gene of wheat(Triticum aestivum L.)[J]. Journal of Triticeae Crops,38(11):1300-1308.]
相吉山,穆培源,桑伟,聂迎彬,徐红军,庄丽,崔凤娟,韩新年,邹波. 2014. 小麦粒重基因TaCwi-A1功能标记CWI22、CWI21的验证及应用[J]. 中国农业科学,47(13):2671-2709. [Xiang J S,Mu P Y,Sang W,Nie Y B,Xu H J,Zhuang L,Cui F J,Han X N,Zou B. 2014. Validation and application of function markers CWI22 and CWI21 of Tacwi-A1 gene related to wheat kernel weight[J]. Scientia Agricultura Sinica,47(13):2671-2709.]
英敏,余虎. 2012. “十一五”贵州小麦区试结果分析与评价[J]. 种子,(5):101-102. [Ying M,Yu H. 2012. Analysis and evaluation on wheat regional trial in Guizhou Pro-vince during ‘11 th five-year plan[J]. Seed,(5):101-102.]
Dong Y M,Jun Y,Zhong H H,Ling W,Xian C X. 2012. Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers[J]. Molecular Breeding,29:43-52.
Hunter M C,Smith R G,Schipanski M E,Atwood L W,Mortensen D A. 2017. Agriculture in 2050:Recalibrating targets for sustainable intensification[J]. Bioscience,67(4):385-390.
Jiang Q Y,Hou J,Hao C Y,Wang L F,Ge H M,Dong Y S,Zhang X Y. 2011. The wheat(T. aestivum) sucrose synthase 2 gene(TaSus2) active in endosperm development is associated with yield traits[J]. Functional & Integrative Genomics,11:49-61.
Ma D Y,Yan J,He Z H,Wu L,Xia X C. 2012. Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers[J]. Molecular Breeding,29:43-52.
Su Z Q,Hao C Y,Wang L F,Dong Y C,Zhang X Y. 2011. Identification and development of a functional marker of TaGW2 associated with grain weight in bread wheat(Triti-cum aestivum L.)[J]. Theoretical & Applied Genetics,122(1):211-223.
Tian J C,Deng Z Y,Hu R B,Wang Y X. 2006. Yield components of super wheat cultivars with different types and the path coeffificient analysis on grain yield[J]. Acta Agro-nomica Sinica,32(11):1699-1705.
(責任编辑 陈 燕)
猜你喜欢
安徽农业科学(2019年14期)2019-08-27
现代农业研究(2019年3期)2019-06-05
吉林农业(2017年7期)2017-07-12
江苏农业科学(2016年8期)2017-02-15
山东农业科学(2016年12期)2017-01-21
天津农业科学(2016年12期)2017-01-11
职工法律天地·下半月(2016年10期)2016-11-30
资治文摘(2016年7期)2016-11-23
农业与技术(2016年15期)2016-11-09
企业导报(2016年8期)2016-05-31
