
猪精液冷冻保存技术研究进展
2020-11-25 02:13谢言射徐志谦蔡更元吴珍芳洪林君黄思秀
家畜生态学报 2020年10期
谢言射,徐志谦,蔡更元,吴珍芳,洪林君,黄思秀
(华南农业大学 动物科学学院/国家生猪种业工程技术研究中心,广东 广州 510642)
猪精液冷冻保存技术是利用液氮(-196 ℃)做为冷源,在超低温环境下冷冻处理猪精液,完全抑制其精子的生理代谢活动,达到长期保存猪精液的目的。冷冻保存技术对保存优质遗传资源、提高种公猪利用率、防止疾病传播、推动联和育种、加速中国生猪产业转型等方面具有重大作用。然而,中国猪精液冷冻保存技术起步较晚,多以引进国外相关设备和技术为基础,目前虽然在精液稀释液、冷冻保护剂、冷冻程序等方面的研究和优化中取得了一定成绩,但精液冷冻保存、冷冻损伤等的机理尚未完全清楚,仍需要进一步探究。本文结合猪精液冷冻保存技术的发展简况、影响猪精液冷冻过程精子质量的因素、猪精液稀释液的机理,对精液冷冻保存的最新研究进展进行了综述,为相关研究以及生产实际应用提供理论参考。
1 猪精液冷冻保存技术的发展简况
精子是第一个成功冷冻保存的哺乳动物细胞。1949年Polge等[1]在试验中偶然发现甘油对冷冻保存的影响,并通过后续试验发现含有20%甘油的精液稀释液能使在-80 ℃下短暂冷冻的家禽精液完全恢复活力,这标志着血细胞、精子以及其他组织、器官冷冻保存方法的开始。精液的第一次冷冻保存也为低温生物学理论的研究提供了动力,尤其是旨在发现细胞在低温条件下受损或死亡原因的研究。1956年,Polge等[2]对不同哺乳动物的精液进行冷冻保存,发现不同哺乳动物精液冷冻保存的最适甘油浓度不同,其中7.5%甘油的稀释液对猪精液的毒性最小,猪精液在解冻后仍存在活性但没有受精能力。1970年以后,猪精液冷冻技术发展较快。1975年Pursel等[3]用颗粒冷冻法冷冻保存猪精液,使用解冻后精液对26头母猪进行人工授精,其中22头母猪受孕且19头母猪的受精卵发育正常。 1975年Westendorf等[4]用细管冷冻精液对30头青年母猪和经产母猪进行人工授精,受胎率高达70%。此后,各国研究人员对不同方面的猪精液冷冻保存技术进行研究,包括稀释液配方、各种冷冻保护剂的选择、冷冻程序的优化、冷冻损伤机理等。而中国猪精液冷冻保存技术研究起步较晚,直到1974 年才由广西壮族自治区畜牧研究所首先开展研究,1980年以后开始从国外引进生产冷冻精液的设备和技术,精液冷冻保存技术进入快速发展阶段。1978年11月至1980年12月,王丕建等[5]将葡萄糖4 g、蔗糖1.2 g、新鲜牛奶30 mL、新鲜蛋黄10 mL、甘油3 mL、蒸馏水57 mL配成的稀释液按1:1比例稀释猪精液,解冻后人工授精132头母猪,情期受胎率51.92%,平均每窝产仔数10.19头。1991年滕钟秀等[6]通过分析精子活率、精子复苏率、顶体完整率、乳酸脱氢酶活性和谷草转氨酶活性等5个方面得出含2%甘油的精液稀释液保存效果最佳。自1990起,虽然许多研究人员在精液稀释液、冷冻保护剂、冷冻程序等方面的研究和优化中取得了一定成绩,但对冷冻保存、冷冻损伤等的机理尚未完全清楚,仍需要进一步的探究。
2 影响猪冷冻精液质量的因素
2.1 冷冻环境对精子的影响
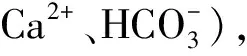
2.2 冷冻过程对精子的氧化应激
精子内存在着复杂的氧化还原体系,通过维持精子内抗氧化物质和氧化物质之间的相互作用,从而影响精子中脂质过氧化反应的水平。精液冷冻保存过程中,精子中存在的某些有效物质失活,打破了抗氧化物质和氧化物质之间的平衡状态,从而产生大量的活性氧物质(reactive oxygen species,ROS),这是影响精液长期冷冻保存的一个重要因素。而ROS在某些水平上又是细胞信号、线粒体功能和免疫等过程所必需的[18],例如猪精子主要能量来源的线粒体氧化磷酸化(果糖)过程中[19],还原性谷胱甘肽参与精子果糖的分解[20],碱性磷酸酶参与精子内果糖的合成与激活[21]。但随着保存过程的进行,用于果糖激活的抗氧化酶的作用和氧化磷酸化的电子传递链将促进ROS的产生。当ROS含量超过生物固有的抗氧化能力时就会产生氧化应激,由于猪精子细胞膜中多不饱和脂肪酸含量高,抗氧化能力有限,所以其对氧化应激更为敏感[22-23]。这些ROS通过精子膜上磷脂的氧化反应改变膜的功能结构,从而破坏精子的功能。同时,ROS还能破坏精子DNA,导致精子受精能力被削弱,从而影响精液体外保存效果。此外,ROS可抑制酶类活性或使酶失活,使ROS不能被及时清除,氧化损伤加深,甚至导致精子凋亡。而且随着精液保存的进行,脂质在自由基的作用下发生过氧化反应,氧化终产物为丙二醛(Malondialdehyde ,MDA),MDA水平随着精液保存时间的增加逐渐升高,并通过与蛋白质的交联作用进一步损害精子细胞的功能[24-25]。
2.3 冷冻速率对精子的影响
精子的损伤程度与冷冻速率有关,猪精子的最佳冷冻速率为0.1 ℃/min[26],当冷冻速率过低(缓慢冷冻损伤)或冷冻速率过高(快速冷冻损伤)时细胞存活率降低[27]。当细胞缓慢冷冻时,细胞脱水最为严重,细胞内的盐浓度最大,对细胞本身造成巨大的损害。当冷冻速率增加时细胞内脱水减少,细胞内溶质浓度减少,细胞收缩减少。然而,当冷冻速率过快时,脱水速度可能不够快,无法防止细胞内的冰核形成[27-29],产生冰核作用,引起一系列损害。一般来说根据含水量、细胞的大小和形状以及胞质膜的透水系数,每个细胞都将有一个特定的临界冷冻速率范围,这意味着对于单个动物的精子来说,最佳冷冻速率应该是确定的,但不同的公猪在相同冷冻速率处理后的精子存活率有很大差异。2005年,Holt等[30]通过对大白、长白、杜洛克三个品种共129头公猪进行试验,发现在相同冷冻速率下长白猪的精子解冻后质量最好,为了获得更准确的结果,Holt等用扩增片段长度多态性技术对解冻后精子质量好的公猪DNA和解冻后精子质量差的公猪DNA进行比较,发现与解冻后精子质量有关的DNA图谱中存在16条有显著差异的条带,表明公猪个体之间的基因组差异与解冻后精子质量相关,这意味着仍存在研究评估其他更快冷冻速率的可行性。
2.4 其他影响猪精液冷冻保存的因素
2.4.1 酸碱度对精子质量的影响 射精后,精子的酸碱度(pH)约为7.4[31]。随着冷冻的进行,细胞脱水,某些细胞内溶质浓度上升达到饱和,产生结晶。这种选择性沉淀将引起pH的下降,并通过损伤蛋白质或改变脂质双层结构从而严重影响膜的稳定性。除此之外,低的pH将引起精子凝集,甚至激活pH依赖的蛋白水解酶,如半胱天冬氨酸蛋白酶,能够导致细胞过早凋亡[32]。此外,外部酸碱度可能影响精子的鞭毛运动,1993年Gatti等[33]通过采集精液并在不同pH培养基中培养,发现射精后精子在精浆中具有高度的运动性,在pH5的培养基中培养5 min,精子运动率仅为5%;在pH7的培养基中培养5 min,精子运动率超过70%;在pH7~9的培养基中培养5 min,所有运动精子都呈前向运动,鞭毛搏动频率平均为15~20 Hz。随着pH的下降,鞭毛搏动频率下降且越来越不规律。
2.4.2 细菌对精子质量的影响 健康公猪的精液基本上都会被细菌感染,细菌含量高达109个/mL。这些细菌部分属于内源性污染,公猪的包皮、尿道、外生殖器及泌尿生殖感染等因素均可能导致精液被细菌污染,而多数属于外源性污染,因为采精器具、稀释液及保存环境等人为过程中缺乏相应的卫生条件导致精液被细菌污染,尤其在高温季节病原菌易滋生。但在精液中检测到的大部分细菌都是无致病性的,因为雌性生殖道自身进化出了一种对抗精液中细菌的防御机制,所以在自然交配的情况下,生殖性能几乎没有受到影响。然而,当使用人工授精时,细菌能在稀释后的精液中生长繁殖,这些精液在雌性生殖道沉积后可能导致胚胎或胎儿死亡、子宫内膜炎、全身感染等问题[34]。冷冻保存过程中细菌等病原菌对猪精子质量产生的较大影响主要体现在诱导精子发生凝集现象[35],降低精子运动参数等质量特性,诱发破坏精子顶体完整性,抑制精子特异性蛋白质磷酸化水平[36],降低精液 pH 以及母猪产仔数等生产性能[37]。除此之外,有研究强调精液稀释液中的抗生素对精液中细菌的抑制作用有限[38],为了避免这些问题,在采集过程和冷冻保存过程中应当遵守相应的卫生守则。
2.4.3 精液内某些离子对精子质量的影响 精浆内钙浓度与前向运动精子比例呈负相关性,因为钙能够调节精子活性。这种调节是由钙浓度决定的,低浓度时能提升精子活性,高浓度时抑制精子活性。铁和硒对质膜完整性呈正相关性[39]。冷冻保存的猪精液在有氧环境下进行活跃的新陈代谢,为维持氧化作用和抗氧化作用的平衡,防止氧化作用带来损伤,铁被铁依赖酶(过氧化氢酶)消耗,硒被用于控制谷胱甘肽过氧化酶水平而消耗。因此,这些金属的含量将在冷冻保存过程中下降,降低抗氧化能力。
3 猪精液稀释液的机理
目前有多种商用精液稀释液用于保存猪精子,提高猪冷冻精液解冻后活力和受精能力,主要由冷冻保护剂、抗氧化物质、营养物质[40]、抗冷休克物质、缓冲剂、无机盐[31]、抗生素[41]组成。
冷冻保护剂是猪精液稀释液的重要组成成分,可分为渗透性和非渗透性冷冻保护剂。渗透性冷冻保护剂相对分子质量较小,容易透过细胞膜到达细胞内部,主要包括甘油、乙二醇、丙二醇、 甲醇等,甘油是目前应用最广泛的冷冻保护剂之一。甘油的加入能够增加其他溶质(甘油和其他盐)的浓度并停止冰的形成,到达比没有甘油时更低的盐浓度,从而使未冻结的部分变大,使细胞的收缩程度变小。除此之外,甘油可以与膜脂的极性头基团形成氢键[15,42]。在正常的水合物细胞中,极性头基团将由水分子(氢键)水合,这对膜的稳定性极为重要[15]。因此,甘油在一定程度上具有在脱水条件下取代水的能力,增加膜的稳定性。而且甘油能够改变保护剂的机械性能,甘油粘度增加可以防止溶质共生结晶[43]。而且随着粘度增大,促进细胞玻璃化形成,改变未冻溶液通道的形状,增加未冻溶液的塑性,减少机械损害。非渗透性冷冻保护剂的相对分子量一般较大,主要以单糖、双糖为主,所以在冷冻保存过程中不能渗入细胞内,主要通过增加细胞外液渗透压,促使细胞在冷冻时脱水,减少胞内冰晶形成,从而保护精子免受机械损伤[44]。目前,海藻糖引起了研究人员的高度关注。海藻糖是一种十分稳定的非还原性双糖,与蔗糖和麦芽糖等具有相似的分子结构和生理作用,是天然双糖中最稳定的糖类,而且对细胞无毒害作用。海藻糖具有独特的非特异性保护作用,可以保护细胞磷脂的双分子层,在冷冻过程中通过细胞膜外产生一种玻璃状的保护层,提高细胞对胞外高渗溶液的耐受性,从而保护精子活性[45]。
4 其他相关研究
4.1 冷冻保存前的保温时间
冷冻保存公司基本上远离猪场。为了进行运输,一般将精子在精液稀释液中稀释(例如BTS)并在15~17 ℃保存。这一时期就叫做保温时间,除了能使猪精液的冷冻保存更加便利外还能提高精子的抗冻性。但理想的保温时间并不确定,Torres等[46]通过分析不同冷冻保存前的保温时间(0, 4, 8, 12, 24, 28 和 32 h)对解冻后精子质量的影响,从精子动力学、质膜和顶体膜的完整性、线粒体膜电位、超氧化阴离子检测以及质膜流动性和过氧化作用等方面进行分析,得出在冷冻保存前使用24 h的保温时间能够增加运动精子、前向运动精子、快速精子的比例,增加质膜、顶体膜的完整性。至于线粒体膜电位,32 h是效果最好的保温时间。但是过氧化阴离子等级和脂质过氧化程度会随时间的增加而增加,和线粒体膜电位增加相比,过氧化阴离子等级和脂质过氧化程度增加弊大于利。而保温时间对质膜流动性影响不大。因此,为了得到最好精子品质,理想的保温时间是24 h。
4.2 烟酸对质膜功能及不饱和脂肪酸组成的影响
烟酸(nicotinic acid,NA)是维生素B3的衍生物,属于维生素B家族,它不但有助于产生能量,还用于脂肪酸和类固醇的合成。而且NA的抗氧化能力、细胞膜保护能力以及其他作用都已经被大量报道。在以往的研究中,NA不但提高了精子存活率和顶体完整性,而且提高了线粒体活性,抑制了顶体反应,提高了解冻后精子的质量。此外,NA具有稳定的自由基、活性氧和细胞膜保护能力,但对猪精子冷冻过程中质膜功能和多不饱和脂肪酸组成的影响尚不清楚。因此,Lee等[47]研究了不同浓度NA对解冻后精子质膜结构和渗透平衡、脂质过氧化以及饱和脂肪酸和多不饱和脂肪酸组分比值相关的生理特性的影响。结果显示,10 mmol/L NA处理的解冻后精子质膜完整性明显高于0 mmol/L和20 mmol/L NA处理组,且脂质过氧化程度明显下降,饱和脂肪酸显著降低,多不饱和脂肪酸显著增加。总之,10 mmol/L的NA改善了质膜的完整性,抑制了脂质过氧化,并增加了解冻后精子的多不饱和脂肪酸含量。这些结果表明,在猪精子冷冻保存中,NA对保护质膜和抑制多不饱和脂肪酸的损耗可能是有用的。
4.3 精浆分离对冷冻保存后精子特性的影响
目前包括冷冻猪精液不同组分的多种方法正被用于改善冷冻保存猪精子质量。猪精液是一种复杂的混合物,包含大量与各种受精有关的蛋白质成分。虽然大多数猪精浆蛋白在精子功能中的作用尚未完全了解,但有证据表明精液组分对解冻后精子质量有影响。为了探究精浆分离对猪冷冻后精子质量的影响。Wasilewska-Sakowska等[48]通过凝胶过滤色谱法得到两个精浆组分:精浆1(>40 kDa)和精浆2(<40 kDa)。解冻前用分离的精浆(精浆1和精浆2)、全精浆和BTS解冻液处理精液。结果显示,全精浆处理的精子质量明显低于分离精浆处理和BTS处理的精子质量;分离精浆处理的精子解冻后顶体完整性和活力明显高于BTS处理的精子。但是经精浆1和精浆2处理的精液样品解冻后精子质量并没有差异。然而,不同试验组精子蛋白的电泳图谱却存在显著差异,说明猪精液不同组分的低温保护作用不同,分离后的精浆对解冻后精子质量有不同影响。此外,作者发现色谱分离法可用于解决分离精浆过程中高丰度精浆组分使低丰度精浆组分无法被识别的问题。
4 总结及展望
猪精液的冷冻保存技术是推动联合育种、加速中国生猪产业转型的重要步骤,也对保存优质遗传资源、提高种公猪利用率、防止疾病传播等有着重要意义。然而,解冻后精子活力、受胎率和窝产仔数方面与鲜精相比仍存在较大差距。此外,公猪廉价的经济价值无法匹配昂贵的仪器设备和复杂的冷冻程度等问题,导致猪精液的冷冻保存技术无法在实际生产中推广。猪精液冷冻保存技术的关键在于减少精子在冷冻过程中的损伤,提高解冻后精子的活性和生殖能力,对冷冻程序进行优化。尽管近年来精液冷冻保存技术在精液稀释液、解冻剂、冷冻程序等方面进行了深入探索并取得了一定进展,但精液冷冻保存机理尚未完全清楚。因此,就目前冷冻保存技术存在的问题来看,未来的研究可分为以下几个方面:一是建立一套更加完善的猪精液冷冻保存体系并研发更适用于实际生产的精液冷冻保存设备;二是结合其他学科,累积研究经验,从多角度研究猪精液冷冻保存技术;三是深入研究猪精液冷冻保存机理,力求从新的方面研究猪精液冷冻保存技术。
猜你喜欢
辽宁大学学报(自然科学版)(2022年1期)2022-04-26
中国饲料(2021年17期)2021-11-02
中国畜牧杂志(2021年3期)2021-03-15
装备环境工程(2021年2期)2021-03-08
中国电气工程学报(2020年5期)2020-10-27
中学科技(2017年11期)2017-12-26
农村农业农民·B版(2017年4期)2017-05-13
中学生数理化·八年级物理人教版(2015年10期)2016-01-04
云南畜牧兽医(2012年5期)2012-11-14
山东农业科学(2009年7期)2009-12-31
