
膜荚黄芪香豆酸辅酶A连接酶基因(4CL)的克隆与表达分析
2020-11-23 07:53冯艺川吴松权全雪丽
延边大学农学学报 2020年3期
赵 洋, 冯艺川, 哈 洋, 吴松权, 全雪丽
(延边大学农学院,吉林 延吉 133000)
膜荚黄芪(Astragalusmembranaceus(Fisch.)Bge.)为豆科多年生草本植物,国家三级濒危保护植物,具有补气升阳、生津养血、行滞通痹等功效[1]。据已有文献报道,膜荚黄芪中的主要药用活性成分为黄酮类、皂苷类等化合物[2-5]。《中华人民共和国药典(第一部)》[1]规定毛蕊异黄酮葡萄糖苷是其主要活性指标成分之一,具有增强机体免疫力、抗炎、降血糖和保护肝脏等作用[6-9]。目前,黄芪毛蕊异黄酮葡萄糖苷的生物合成研究已成为研究热点。
4-香豆酸辅酶A连接酶(4-coumarate:CoA lig-ase,4CL)是植物类黄酮类化合物和木质素等生物合成途径的一种关键酶。据已有文献报道,拟南芥、黑麦草、欧洲山杨、覆盆子、毛白杨、亚麻、青稞、甜高粱等多种植物中已成功克隆出了4CL基因[10-11]。此外,4CL基因既在植物生命世代初期开始由植物自身的发育调控,还能被多种诱导子及环境因子激活[12]。而有关膜荚黄芪4CL全长基因克隆的研究尚未见报道。本研究以膜荚黄芪为材料,通过RACE法克隆了4CL全长cDNA序列,并利用生物信息学和qPCR转录表达方法[13]对其功能进行了分析。
1 材料与方法
1.1 材料
1) 植物材料 2年生膜荚黄芪。选取一部分的根、茎、叶,用液氮冷冻处理后置于-80 ℃的超低温冰箱中,另一部分样品置于50 ℃恒温干燥箱中烘干至恒定后用于成分分析。
2) 试剂及仪器 Trizol试剂,购自Invitrogen公司;合成cDNA第一条链的试剂盒,购自Toyobo公司;SMARTerTMRACE cDNA 5’和3’RACE试剂盒,购自Clontech公司;DNA凝胶回收试剂盒和PMD-19T载体试剂盒,购于宝生物工程(大连)有限公司;对照品毛蕊异黄酮葡萄糖苷(批号110907),购自上海融禾医药科技有限公司;乙腈和甲醇为色谱级,水为超纯水,其他试剂分析纯,购于国内各公司。
1.2 方法
1.2.1 引物设计
在NCBI网站上搜索已经在Genbank上登陆的豆科植物及拟南芥等模式植物的4CL的mRNA和蛋白质的完整或部分序列并下载,使用MegAlign软件确定其保守区域后,利用Primer Premier 5.1软件设计1对Am4CL基因的简并引物Am4CLu1和Am4CLd1(引物的合成由上海英骏生物合成公司完成)。该试验所用引物如表1所示。

表1 引物序列
1.2.2 总RNA提取与cDNA的合成
采用Invitrogen公司的Trizol一步法提取膜荚黄芪根总RNA,具体步骤参照其说明书,将提取的总RNA保存于-80 ℃超低温冰箱中。在150 V电压、1.2%琼脂糖凝胶电泳15 min和NanoDrop 2 000 C超微量分光光度计检测其质量与浓度;按照TOYOBO公司的ReverTra Ace qPCR RT Master Mix with gDNA Remover反转录试剂盒说明书的步骤合成膜荚黄芪cDNA第1条链。
1.2.34CL基因保守区克隆
以合成的cDNA第1条链为模板,进行PCR扩增,反应体系共20.0 μL:10×PCR Buffer 2.0 μL,25 mmol/L MgCl22.0 μL,2 mmol/L dNTP 2.0 μL,cDNA 1 μmol/L 2.0 μL,Am4CLu1 2.0 μL,Am4CLd1 2.0 μL,5 U ExTaq 0.2 μL,dd H2O 7.8 μL。使用快速琼脂糖凝胶DNA回收试剂盒(康为世纪公司)回收PCR产物的DNA片段;将目的片段与pMD18-T载体在16 ℃条件下进行过夜连接;将重组质粒转化至大肠杆菌感受态JM109;将提取的重组质粒DNA原液稀释50倍后,使用M13引物(表1)进行PCR检测;将PCR产物在1.2%的琼脂糖胶上电泳,将阳性克隆菌株进行测序。测序工作由上海英骏生物公司完成。
1.2.4 5’RACE 和 3’RACE 扩增
参照Clontech公司的SMARTerTMRACE cDNA扩增试剂盒说明书,分别合成目的基因片段的5’端和3’端。根据 2个4CL基因家族的保守序列分别设计1对 5’-RACE 和 3’-RACE 的特异引物Am4CLgsp1和Am4CLgsp2,引物序列如表1所示。PCR 产物的克隆与测序,方法同1.2.3
1.2.54CL基因编码区的克隆
依据已经克隆的保守区域的cDNA序列和5’-RACE 和 3’-RACE 扩增的产物序列,设计并合成了编码区特异引物Am4CLcu1和Am4CLcd1,如表1所示。反应条件为94 ℃预变性5 min,94 ℃变性45 s,60 ℃退火45 s,72 ℃延伸 2 min,35个循环;72 ℃延伸7 min。PCR产物的克隆与测序,方法同1.2.3。
1.2.64CL基因的生物信息学分析
运用Lasergene软件包中Protean预测编码蛋白分子量和等电点;利用ProtScale(http://web.Expasy.org/protscale/)软件进行疏水性进行分析;利用Psipred预测4CL蛋白的二级结构;利用TMHMM-2.0在线工具进行4CL蛋白序列跨膜区的预测。利用MEGA7软件进行氨基酸序列比对分析和构建4CL氨基酸序列系统进化树。
1.2.7 膜荚黄芪4CL基因的荧光定量PCR分析
针对获得的Am4CL特异序列,设计1对荧光定量PCR引物(表1),所用内参基因为黄芪18s基因(表1),进行荧光定量PCR。反应体系共20.0 μL,除了SYBR 10.0 μL和ROX 0.5 μL外,其他同1.2.3项。以标准的Am4CL和18S基因拷贝数的对数值为横坐标,以测得的CT值为纵坐标,分别绘制其标准曲线。根据未知样品的CT值,可在各自的标准曲线中得到其样品的拷贝数再求出内参基因18S的相对表达量,每组样品进行3次重复检测。
1.2.8 毛蕊异黄酮葡萄糖苷含量的测定
毛蕊异黄酮葡萄糖苷含量采用HPLC方法[14]进行测定,每组样品的提取和测定操作重复3次。
1.2.9 统计分析
试验数据分析采用SPSS 19.0软件,多重比较采用Duncan新复极差法(P<0.05),相关性分析采用Pearson相关系数。
2 结果与分析
2.1 Am4CL基因全长cDNA克隆
用简并引物进行PCR反应获得了4CL基因cDNA片断(图1A),重组质粒进行PCR鉴定得到长度约为500 bp的4CL条带(图1B),将其进行测序,并将片段命名为:Am4CL-RT。
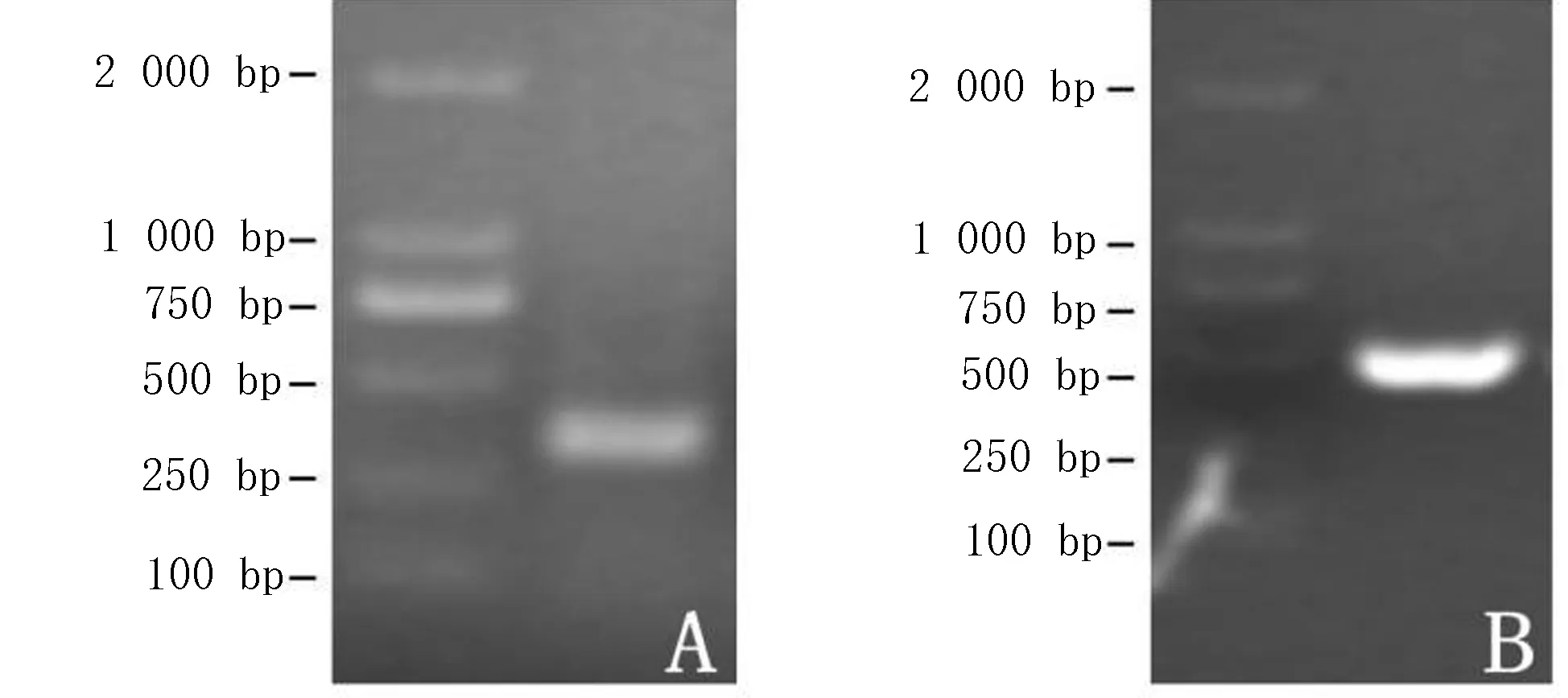
A.用简并引物扩增的4CL基因cDNA片断; B. Am4CL-RT 质粒PCR鉴定
将反转录合成的第1条链,使用设计的特异性引物(表1)分别进行3’-RACE和5’-RACE,电泳如图2(A、C),PCR鉴定如图2(B、D)。Am4CL-3’序列长度为750 bp;Am4CL-5’序列长度为1 300 bp,初步认为Am4CL-3’和Am4CL-5’分别为膜荚黄芪4CL基因的3’端和5’端。
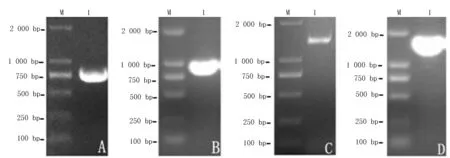
A.Am4CL基因3’端PCR结果;B.Am4CL -3’质粒PCR鉴定;C.Am4CL基因5’端PCR结果;D.Am4CL-5’质粒PCR鉴定
依据所克隆的膜荚黄芪苯丙烷途径基因保守区域序列以及5′-RACE和3′-RACE扩增的产物序列,通过拼接获得目的基因的全长序列并命名为Am4CL,设计编码区序列引物如(表1),以反转录的第1条链cDNA为模板,PCR获得目的基因片段(图3),经过回收、连接、转化、提取重组质粒PCR鉴定。测序结果显示和拼接序列一致。因此,确定获得了膜荚黄芪苯丙烷途径的4CL基因的全长。
2.2 膜荚黄芪4CL基因的序列分析
核苷酸序列和氨基酸序列分析结果表明,Am4CL的cDNA全长为1 937 bp,其中,ORF为1 641 bp,3’非翻译区(3’-UTR)为144 bp,5’非翻译区(5’-UTR)为127 bp,以及25 bp的polyA尾。DNA序列中A+T含量为52.50 %,C+G含量为47.5%,推测其编码546个氨基酸。
登录NCBI主页,用Blast在线搜索工具将Am4CL基因序列与其他植物的4CL基因序列进行对比分析。结果表明,Am4CL豆科的一些植物,比如刺毛黛豆(RDY12997.1)、百脉根(BT139278.1)、红豆(XP_027330824.1)的相似性分别为79%、87%、81%,与豆科其他属的相似性也很高,说明Am4CL是膜荚黄芪的4CL基因序列。
2.3 4CL的生物信息学分析
疏水性分析显示(图4),Am4CL的总平均亲水指数为0.009,是疏水性蛋白质。其中,疏水性最强位点在106位置,达到3.167,亲水性最强位点在192位置,为-2.556。二级结构预测结果(图5),Am4CL蛋白的二级结构α-螺旋占34.07%,β-折叠占18.50%,无规则卷曲占47.44%。16个α-螺旋,19个β-折叠。
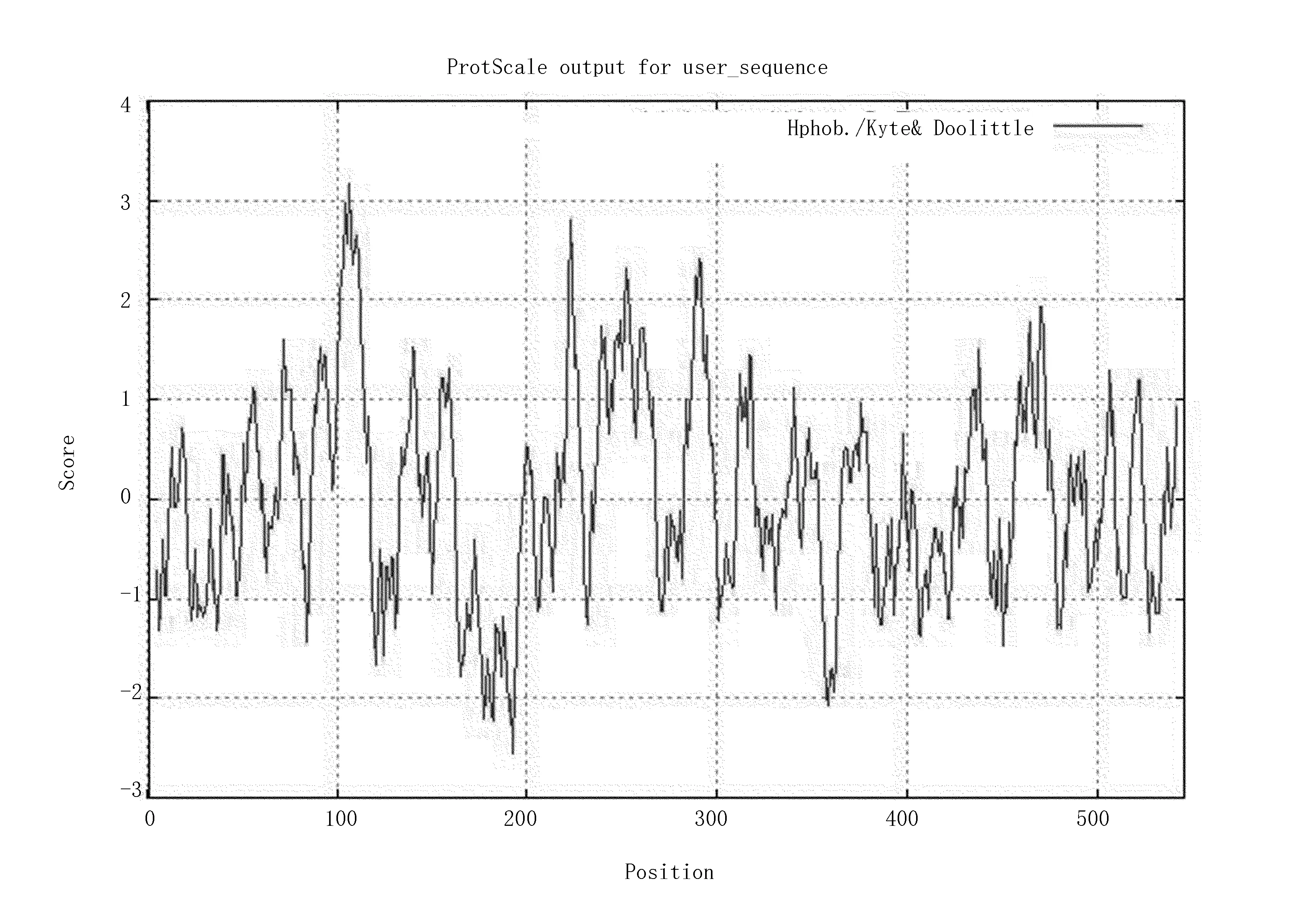
图4 Am4CL蛋白序列的疏水曲线
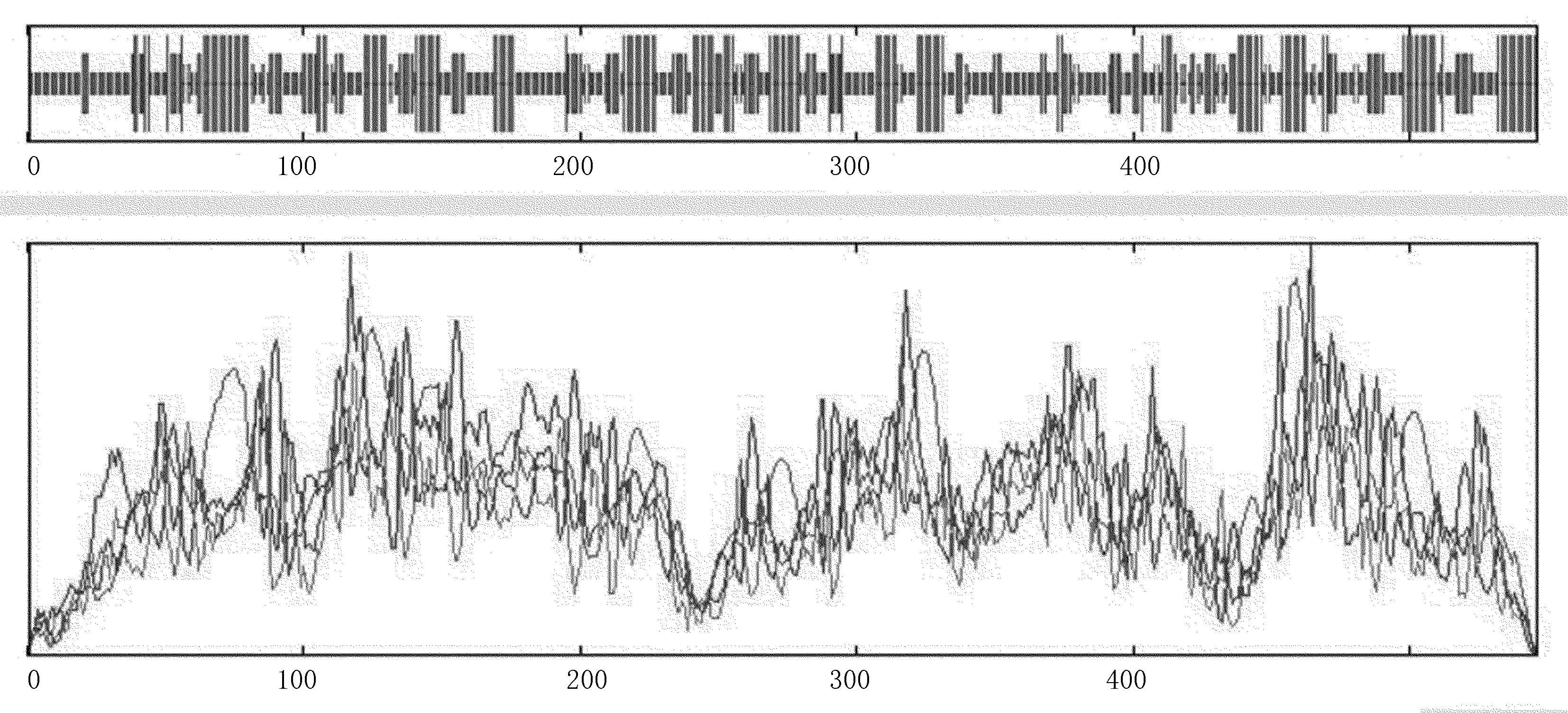
图5 Am4CL编码蛋白的二级结构
用Lasergene软件包中Protean预测Am4CL蛋白相对分子质量为59 470,等电点为8.34,酸性氨基酸48个,碱性氨基酸51个。TMHMM-2.0预测结果显示,Am4CL为膜蛋白,产生了2个跨膜螺旋,其跨膜区预测位置分别位于98~120位氨基酸之间和243~265位氨基酸之间(图6)。
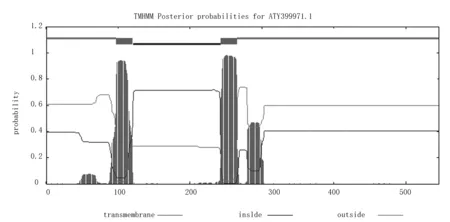
图6 Am4CL蛋白跨膜结构分析Fig.6 Transmembrane structure analysis of protein of Am4CL
为了进一步深入研究Am4CL与其他植物4CL之间的进化关系,以其推测的氨基酸序列作为分析对象,利用DNAstar软件构建系统发育进化树(图7)可以看出14个物种的4CL氨基酸序列被划分为2个主支,分别为TypeⅠ、TypeⅡ。将绿豆变种(XP_014504631.1)、小豆(XP_017430152.1)、菜豆(XP_007160602.1)、大豆(XP_00354004.1)、刺毛黛豆(RDY12997.1)、红豆(XP_027330824.1)、花生(XP_025636299.1)、膜荚黄芪(ATY39971.1)归为Ⅰ类;将紫花苜蓿(AET02374.1、XP_024627525.1、XP_013466211.1)、鹰嘴豆(XP_004499325.1)、地下三叶草(GAU12224.1)、红车轴草(PNY04116.1)归为Ⅱ类。

图7 植物4CL的系统发育进化树
2.4 膜荚黄芪Am4CL基因表达量分析
运用实时荧光定量PCR技术,分析了膜荚黄芪植株根、茎、叶3个不同组织中Am4CL的表达特性(图8)结果表明,Am4CL在根、茎、叶中均表达,但表达量明显存在差异,其中,根中表达量最高,是茎中表达量的2.20倍,是叶中表达量的2.01倍。另测定毛蕊异黄酮葡萄糖苷含量在膜荚黄芪的根、茎、叶中的表达量也存在显著差异,其中,根中含量达到了0.29 mg/g,是茎中含量(0.20 mg/g)的1.45倍,而叶中仅含有0.11 mg/g。
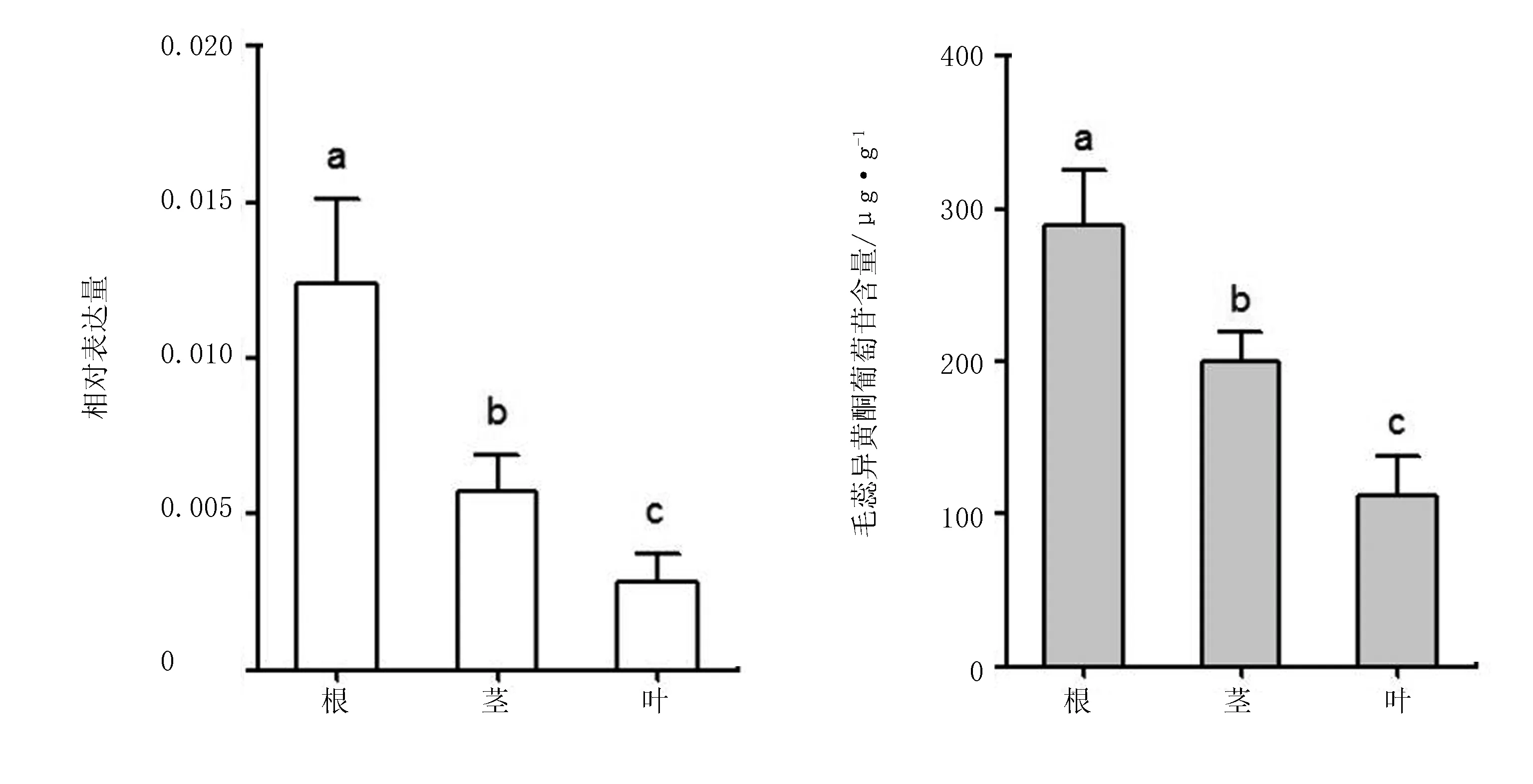
图8 不同组织中4CL基因相对表达量及毛蕊异黄酮葡萄糖苷含量
3 讨论与结论
苯丙烷类代谢途径是植物体次生代谢中的一个通用途径,在植物生长发育中具有重要的作用[15]。4CL是植物苯丙烷类代谢途径的一种关键酶,主要参与木质素和类黄酮合成。Kutsuki等[16]于1981年首次证明,4CL是木质素生物合成途径中的一种酶。冯春燕[17]对几种植物的4CL蛋白比对分析发现,4CL蛋白家族主要与木质素和类黄酮类物质的合成相关,发现大豆的4CL蛋白主要参与了黄酮类物质的合成。罗睿雄等[18]在芒果果实的研究表明,4CL基因可能参与了黄酮类物质的合成,其基因表达与芒果果实着色也具有密切的相关性。Chowdhury等将红麻进行重力胁迫处理,发现与木质素形成的Hc4CL基因有显著上调[19]。本研究借鉴前人的方法和经验,首次从膜荚黄芪中克隆了4CL基因全长序列,用生物信息学软件进行序列比对分析,发现与其他植物中4CL基因有极高的同源性。
运用实时荧光定量 PCR技术对膜荚黄芪中的4CL基因做进一步的研究,在试验过程中发现Am4CL基因在膜荚黄芪的根、茎、叶中均有稳定表达,且根中相对表达量均高于茎和叶,这说明其在根中表达活跃,与白及[20]、丹参[21]等药用植物的研究结果一致,这对以根为药用部位的膜荚黄芪来说具有重要的指示意义。此外还发现,4CL基因在膜荚黄芪的根、茎、叶中的表达趋势与根、茎、叶中的毛蕊异黄酮葡萄糖苷含量趋于一致(图8),证明Am4CL参与毛蕊异黄酮葡萄糖苷的合成。虽然系统进化树(图7)基本按照不同物种分类,但不同物种氨基酸序列的相似性均在77%以上,已有研究表明,豆科植物的4CL蛋白参与了黄酮类物质的合成,推测克隆的Am4CL参与了毛蕊异黄酮葡萄糖苷的生物合成。
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2022年13期)2022-09-15
今日农业(2020年24期)2020-12-15
Digital Chinese Medicine(2020年4期)2020-12-11
食品安全导刊(2020年30期)2020-11-23
海峡姐妹(2019年3期)2019-06-18
中国食品(2018年7期)2018-09-10
家庭医药·快乐养生(2017年4期)2017-04-19
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
