
HNF4A对结直肠癌细胞增殖的影响及其机制*
2020-11-21 03:12吴剑宏曹小年
华中科技大学学报(医学版) 2020年5期
吴 维, 吴剑宏, 曹小年, 杨 熹△
华中科技大学同济医学院附属同济医院 1胃肠外科 3胸外科,武汉 430030 2华中科技大学同济医学院附属武汉市中心医院后湖院区肝胆胰外科,武汉430024
结直肠癌严重影响人们的健康,全世界每年新增超过100万个病例。它的形成涉及基因和表观遗传学改变的多个步骤,包括癌基因、抑癌基因、DNA修复基因以及所有参与结直肠癌发生发展的信号通路[1]。就像大多数实体恶性肿瘤一样,结直肠癌是一种复杂的肿瘤,异形细胞在一个由细胞和非细胞成分构成的复杂微环境中形成了肿瘤[2]。虽然人们长期致力于提高结直肠癌的早期诊断率和探寻有效的治疗方法,但效果并不理想,放化疗或者其他治疗手段有待进一步完善[3]。
肝细胞核因子4(hepatocyte nuclear factor 4,HNF4)有3个亚型:HNF4α,HNF4β,HNF4γ[4]。HNF4α即HNF4A是一种核转录因子,与DNA结合为二聚体。它最初被认为是小鼠肝脏中结合APOC3启动子的一种活性物质[5],随后被认定为青少年的成人起病型1型糖尿病中发生突变的基因[6]。HNF4A的表达具有组织特异性,在肝脏、胰腺、肾脏、肠道的发育及生理功能中不可或缺,尤其是在脂代谢、糖代谢以及炎症反应中起着重要作用[7]。已有文献报道,其编码的蛋白可以作为治疗早期胃癌的潜在靶点[8]。我们前期利用GEPIA生物信息系统分析发现p53的表达与HNF4A正相关。p53基因位于17号染色体,是一个抑癌基因[9]。它可以促进细胞凋亡以及抑制细胞生长,因此它在防止损伤细胞癌变的过程中起着关键作用[10]。有文献表明,长链非编码RNA GClnc1可以通过抑制p53/p21通路来促进结直肠癌的进展[11]。目前,鲜有文章报道HNF4A在肠道中的具体作用,为此,我们探索了HNF4A对结直肠癌生长的影响及可能的机制。
1 材料与方法
1.1 临床资料
选取2016年7月至2017年7月华中科技大学同济医学院附属同济医院收治的结直肠癌患者117例。其中男57例,女60例,年龄40~70岁。取其结直肠癌及癌旁正常上皮组织,提取蛋白,Western blot检测HNF4A蛋白表达量,以高于正常组织HNF4A表达量的结直肠癌患者为HNF4A高表达组,反之则为低表达组,并依据病理结果及影像学资料进行TNM分期。患者纳入标准:①经手术后病理检查或肠镜活检病理诊断为结直肠癌;②年龄≥35岁;③患者或家属均对本研究知情,并自愿签署知情同意书。排除标准:①合并感染性疾病、凝血功能异常等疾病;②已经接受放化疗或免疫治疗;③伴有其他恶性肿瘤;④合并心、肝、脑、肾等重要器官功能障碍;⑤精神严重障碍者;⑥拒绝参与本研究者。
1.2 细胞培养及试剂
结直肠癌细胞系LoVo、SW620、HT-29、SW480均由同济医院分子生物实验室常规保存,于37℃、5%的CO2恒温培养箱中培养。HNF4A、GAPDH、p53、p21一抗均购自美国Santa Cruz Biotechnology公司,兔抗小鼠辣根过氧化物酶标记二抗、山羊抗兔辣根过氧化物酶标记二抗购买于武汉博士德公司。胎牛血清、DMEM培养液、0.25%胰蛋白酶、RIPA细胞裂解液购买于南京凯基公司。Trizol购买于TaKaRa公司。质粒提取试剂盒、转染试剂购自美国Sigma公司。Western blot制胶试剂盒、BrdU细胞增殖检测试剂盒购于武汉启动子公司。HNF4A高表达质粒[载体为pcDNA3.1(+)]及ShHNF4A慢病毒均购于上海吉凯基因公司。引物的合成与核酸的测序由武汉擎科公司完成。
1.3 细胞转染
当LoVo和SW620细胞生长至约50%融合度时进行转染。以无血清DMEM分别稀释Lipofectamine 2000和HNF4A质粒,静置5 min,然后将稀释后的Lipofectamine 2000和HNF4A质粒轻柔混合,静置约20 min。用上述混合液代替原培养液。于培养箱培养6 h后,换回有血清高糖培养液继续培养。
当HT-29和SW480细胞生长至约30%~40%融合度时进行病毒感染。加入108TU/mL ShHNF4A 20 μL,培养48 h后加入1/1000体积嘌呤霉素筛选出稳定低表达HNF4A的细胞。
1.4 Western blot检测蛋白表达
用RIPA裂解细胞提取蛋白并测定蛋白浓度,以SDS-PAGE电泳分离蛋白,然后将蛋白转至PVDF膜,常温封闭1 h,4℃孵育相应一抗过夜(HNF4A、GAPDH、p53、p21抗体以1∶1000稀释),TBST洗膜后,室温下孵育二抗1 h(兔抗及鼠抗以1∶5000稀释),洗膜后曝光显影。
1.5 Real-time PCR检测mRNA表达
于6孔板中正常培养细胞,给予不同处理后待细胞生长至对数期,收集细胞,用Trizol提取细胞RNA,通过Real-time PCR检测并计算各组细胞HNF4A相对表达量。HNF4A上游引物:5′-ATGCGACTCTCCAAAACCCTTCA-3′,下游引物:5′-CTAGATAACTTAATGCTTGGTG-3′;GAPDH上游引物:5′-TGTGGGCATCAATGGATTTGG-3′,下游引物:5′-ACACCATGTATTCCGG-GTCAAT-3′。
1.6 BrdU细胞增殖实验
接种适量细胞于6孔板并进行相应处理,然后用BrdU标记细胞,避光孵育30 min后收集细胞,上流式细胞仪检测。
1.7 MTT法检测HNF4A对细胞增殖的影响
收集各个实验组及对照组细胞,接种于96孔板,密度约104个/孔,依照MTT试剂操作方法,用酶标仪检测其吸光度值(A值),连续测3 d,重复3次。
2 结果
2.1 HNF4A的表达与患者临床特征的关系
收集并分析117例结直肠癌患者病例,并分别按照性别、年龄、淋巴结转移、TNM分期进行分组。结果发现:淋巴结转移患者中高表达HNF4A者明显少于低表达者(P<0.05);肿瘤分期越晚,高表达HNF4A的患者比例越低(P<0.05)。见表1。
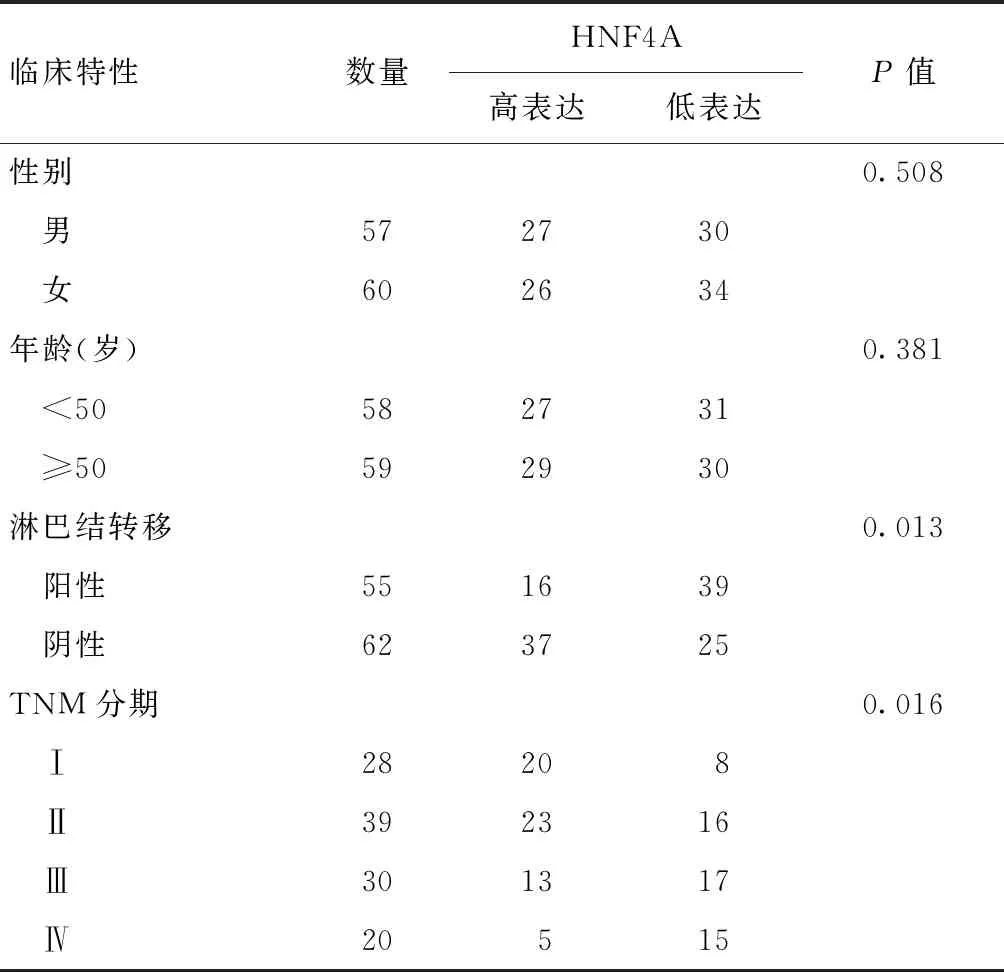
表1 HNF4A的表达量与结直肠癌患者临床特征的关系(例)Table 1 The relationship between the expression level of HNF4A and clinical characteristics(n)
2.2 HNF4A高表达质粒转染LoVo和SW620细胞建立高表达HNF4A细胞系
将HNF4A高表达质粒和相应的对照质粒转染至LoVo和SW620细胞并培养48 h后,收集细胞,分别用Western blot和Real-time PCR检测HNF4A蛋白和mRNA表达水平,可见HNF4A质粒成功转染LoVo和SW620细胞,见图1。
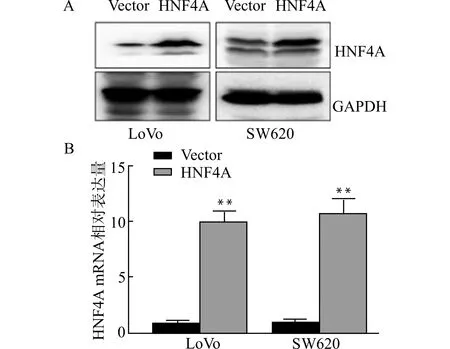
A:Western blot检测HNF4A的蛋白表达量;B:Real-time PCR检测HNF4A的mRNA量;与对照组(Vector)比较,**P<0.01图1 成功构建高表达HNF4A的细胞系Fig.1 Construction of colorectal cancer cell lines with high expression of HNF4A
2.3 转染HNF4A高表达质粒后细胞增殖能力的变化
BrdU标记高表达HNF4A的LoVo、SW620及其相应的对照组细胞,避光孵育30 min后收集细胞,上流式仪检测,结果显示:HNF4A高表达组与对照组细胞相比,细胞增殖能力明显受到抑制(均P<0.01),见图2。
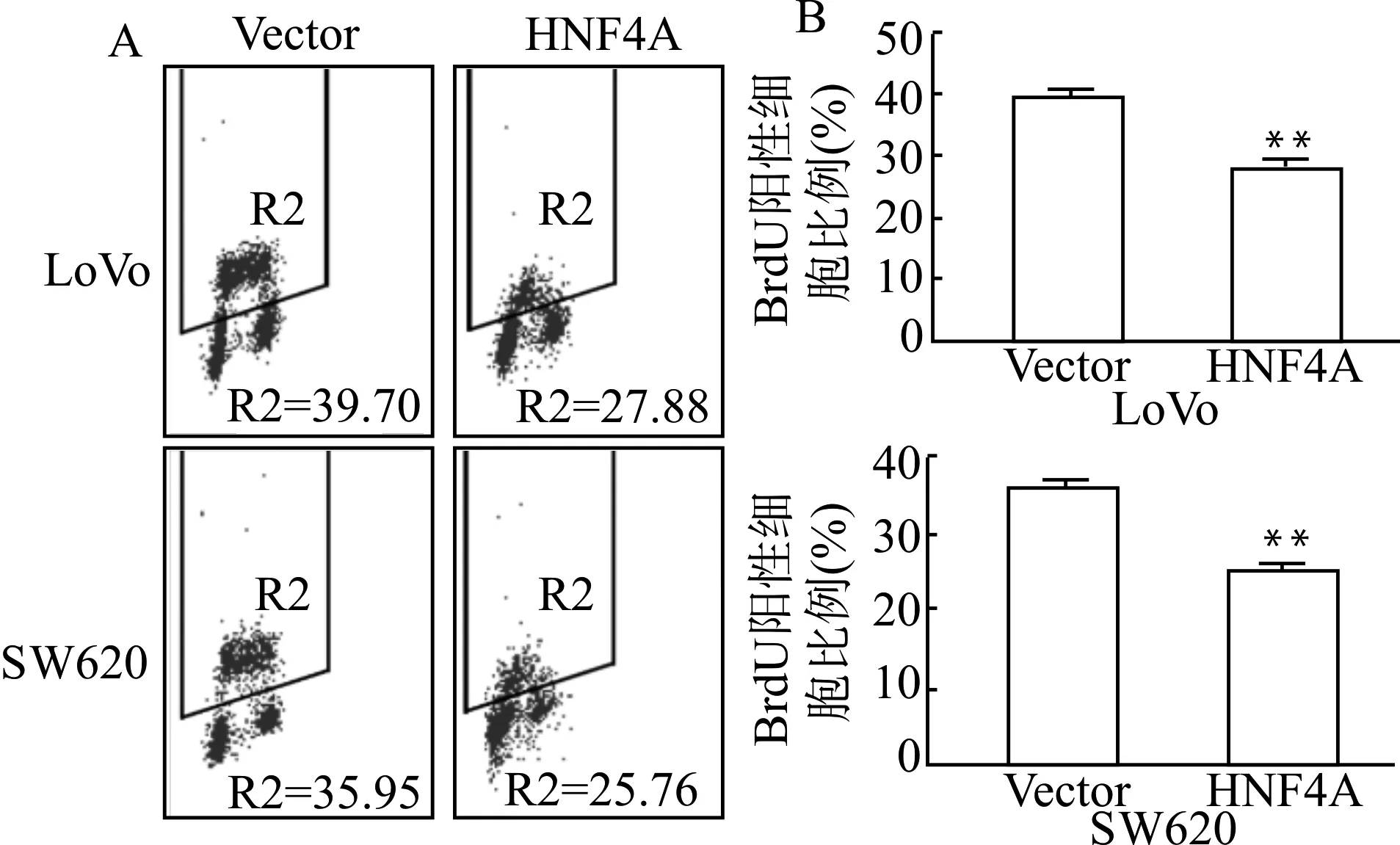
A:流式图;B:统计图;与对照组(Vector)比较,**P<0.01图2 高表达HNF4A对结直肠癌细胞增殖能力的影响Fig.2 Effect of high expression of HNF4A on proliferation of colorectal cancer cells
2.4 慢病毒ShHNF4A转染HT-29和SW480构建稳定低表达HNF4A细胞系
将ShHNF4A#1、ShHNF4A#2以及阴性对照组的慢病毒颗粒转染至HT-29和SW480并培养48 h,加入嘌呤霉素筛选出稳定低表达HNF4A的细胞系,分别用Western blot和Real-time PCR检测HNF4A蛋白表达量和mRNA表达水平,可见慢病毒ShHNF4A成功转染HT-29和SW480细胞,见图3。
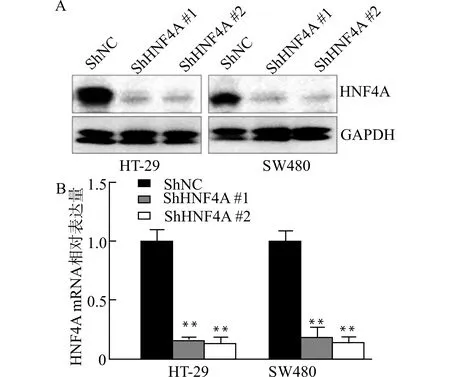
A:Western blot检测HNF4A的蛋白表达量;B:Real-time PCR检测HNF4A的mRNA量;与对照组(ShNC)比较,**P<0.01图3 成功构建低表达HNF4A的细胞系Fig.3 Construction of colorectal cancercell lines with knock-down of HNF4A expression
2.5 转染ShHNF4A慢病毒后细胞增殖能力的变化
通过BrdU细胞增殖实验,我们发现ShHNF4A转染组与阴性对照组(ShNC)比较,细胞增殖能力明显提高(均P<0.01),见图4。
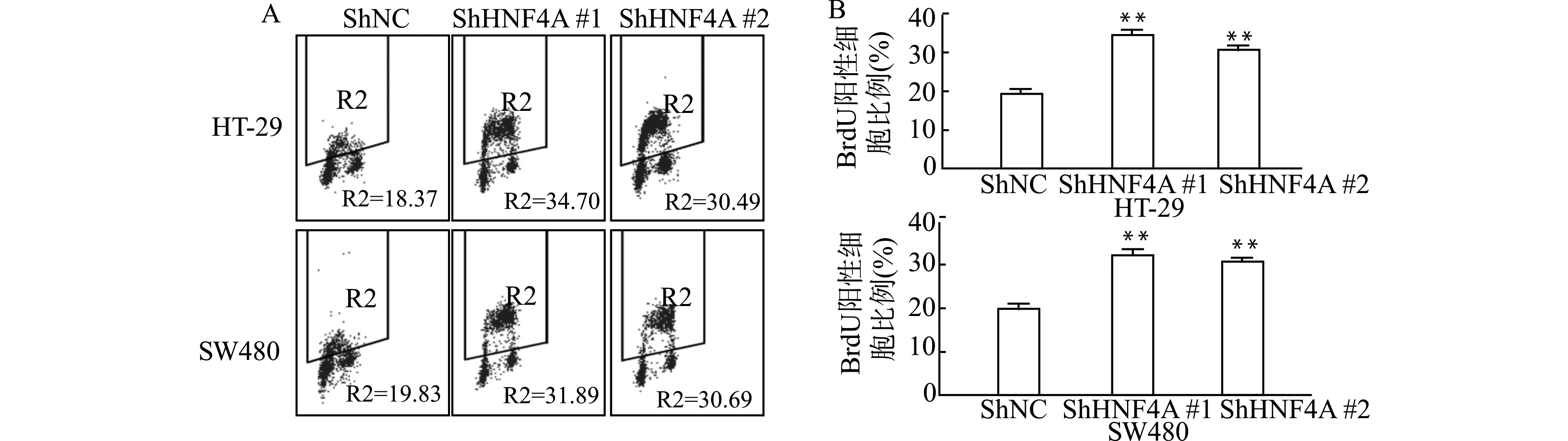
A:流式图;B:统计图;与对照组(ShNC)比较,**P<0.01图4 低表达HNF4A对结直肠癌细胞增殖能力的影响Fig.4 The effect of low expression of HNF4A on proliferation of colorectal cancer cells
2.6 MTT法检测改变HNF4A表达后结直肠癌细胞增殖能力的变化
收集各个实验组及相应对照组细胞接种于96孔板,并设置复孔,以MTT试剂进行增殖实验,酶标仪测定吸光度值。结果提示:在培养72 h时,各组高表达HNF4A的细胞增殖明显受到抑制,而低表达HNF4A的细胞增殖明显增强(均P<0.05),见图5。
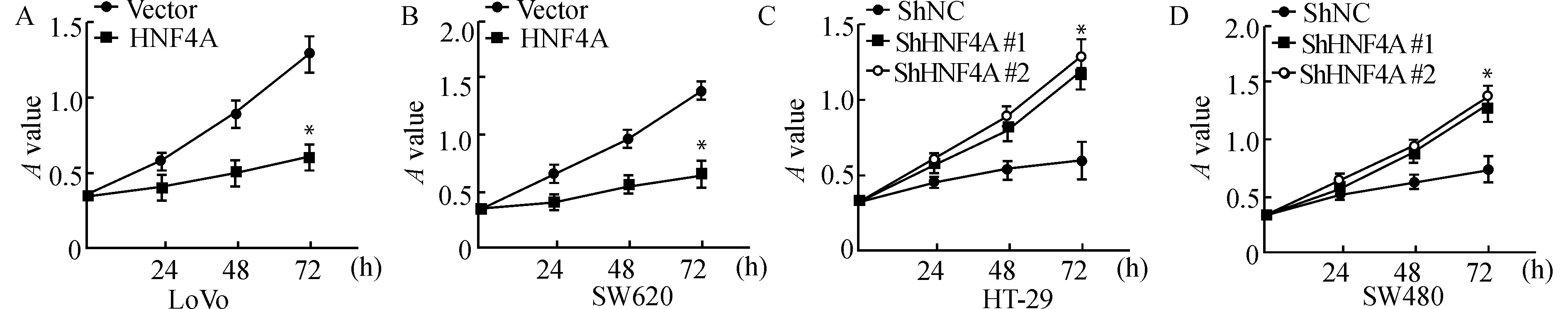
A,B:在LoVo和SW620中高表达HNF4A;C,D:在HT-29和SW480中低表达HNF4A;与对照组比较,*P<0.05图5 MTT实验检测HNF4A表达水平对结直肠癌细胞增殖的影响Fig.5 Effect of HNF4A on cell proliferation verified by MTT assay
2.7 HNF4A通过p53/p21通路影响细胞增殖
p53作为抑癌基因被激活后可以诱导多种基因(包括p21)的转录。p21可以抑制细胞从G1期进入S期,导致细胞增殖阻滞。因此,我们推测HNF4A通过p53/p21信号通路发挥作用。Western blot实验证明高表达HNF4A可以诱导p53的表达,进而促进p21表达;反之,低表达HNF4A则抑制p53及p21的表达,见图6。
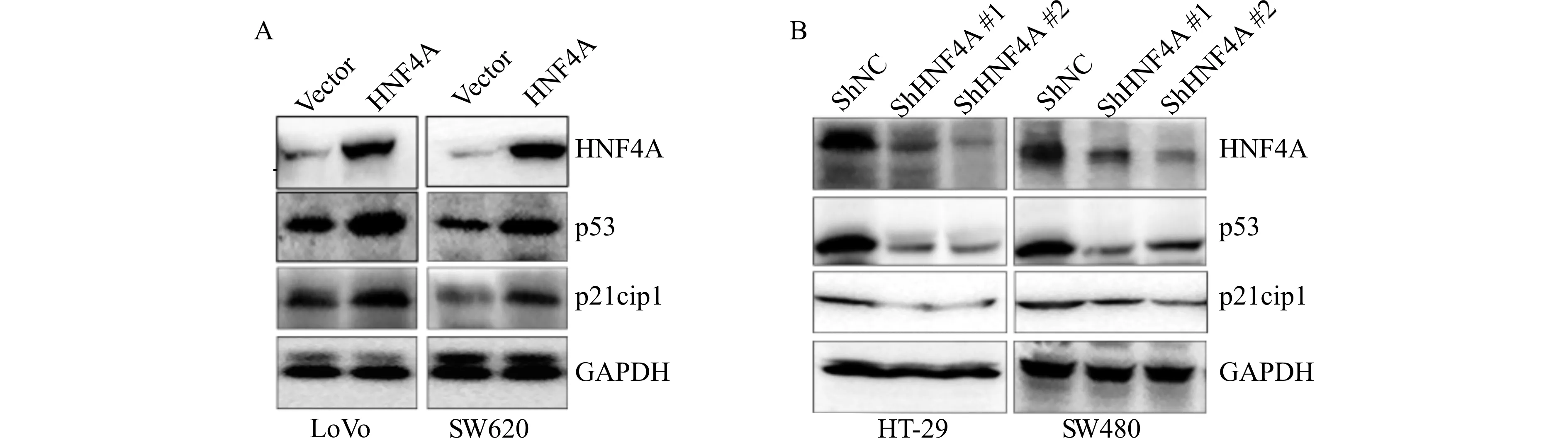
A:在LoVo和SW620中高表达HNF4A后,通过Western blot检测p53及p21蛋白表达量;B:在HT-29和SW480中低表达HNF4A后,通过Western blot检测p53及p21蛋白表达量图6 Western blot检测改变HNF4A表达水平后p53及p21的蛋白表达变化Fig.6 The expression levels of p53 and p21 detected by Western blotting after high and low expression of HNF4A
3 讨论
虽然近些年治疗策略得到了改进,但结直肠癌仍是全世界范围内因癌症死亡数量最多的肿瘤类型[12]。随着对结直肠肿瘤分子机制的不断认知,已经有相关的分子靶向治疗可以有效提高临床治疗效果,使转移癌的中期生存时间从6~10个月提高到30个月甚至更长[13]。
有研究表明,在肝细胞癌中,HNF4A是调控上皮间质转化(EMT)的重要因子[4]。它也可以控制肝脏上皮细胞的形成、肝脏的形态发育,以及抑制肝癌的发生。此外,有学者发现在肾细胞癌中,HNF4A呈高表达并且非常活跃,可以通过调控其下游信号通路改变肾细胞癌的细胞形态[14]。有文献报道,HNF4A可以通过Wnt/β-catenin信号通路抑制结直肠癌的恶化[15]。另有学者指出,在结肠炎及结直肠癌中,HNF4A的亚型在特定的情况下发挥对立的作用[16]。本研究从结直肠癌患者的临床病理资料开始,分析了HNF4A的表达与结直肠癌的可能关系,进而在不同的结直肠癌细胞中验证发现,HNF4A与结直肠癌细胞的增殖呈负向调节关系,这为HNF4A成为结直肠癌的治疗靶点提供了理论基础。
肿瘤抑制因子p53可以影响细胞凋亡和细胞周期,而在这些过程中,Bax和p21是重要的介质[17]。p21有p53依赖型和非p53依赖型两种调节方式,但p53依赖型p21调节对于细胞周期阻滞至关重要[18]。p53是p21的主要调节因子,当细胞接受例如DNA损伤等一系列刺激时,可以诱导p21的表达[19]。有文献报道,RASSF1A通过激活p53依赖的p21途径抑制结直肠癌细胞的生长[20]。在所有恶性肿瘤中,50%以上会出现p53的突变。p53基因突变后,由于其空间构象发生改变,失去了对细胞生长、凋亡和DNA修复的调控作用,由抑癌基因转变为癌基因。因此,我们推测HNF4A可能对p53的表达产生影响,并且我们在数据库中发现HNF4A与p53的表达存在正相关趋势。
因此,本研究检测了高、低表达HNF4A之后p53和p21蛋白的表达变化,结果发现:高表达HNF4A后,p53和p21的表达量也升高;反之,低表达HNF4A后,p53和p21表达量降低。这项结果表明HNF4A在结直肠癌的发生发展中起着重要的调控作用,我们在今后的研究中将进一步探索其具体的调控机制。
猜你喜欢
住区(2022年5期)2022-12-30
美文(2022年4期)2022-03-07
科学与生活(2021年16期)2021-11-25
高考金刊·理科版(2019年8期)2019-02-14
医药前沿(2019年2期)2019-01-05
投资者报(2018年37期)2018-09-19
医学研究杂志(2015年9期)2015-07-01
医学研究杂志(2015年9期)2015-07-01
癌变·畸变·突变(2015年4期)2015-02-27
祝您健康(1989年2期)1989-12-30
