
箭叶淫羊藿与近缘种天平山淫羊藿花粉形态对比研究
2020-11-02 22:23:05黄华赖华清蒋勇李仁清李风琴刘勇徐艳琴
广西植物 2020年9期
关键词:居群
黄华 赖华清 蒋勇 李仁清 李风琴 刘勇 徐艳琴
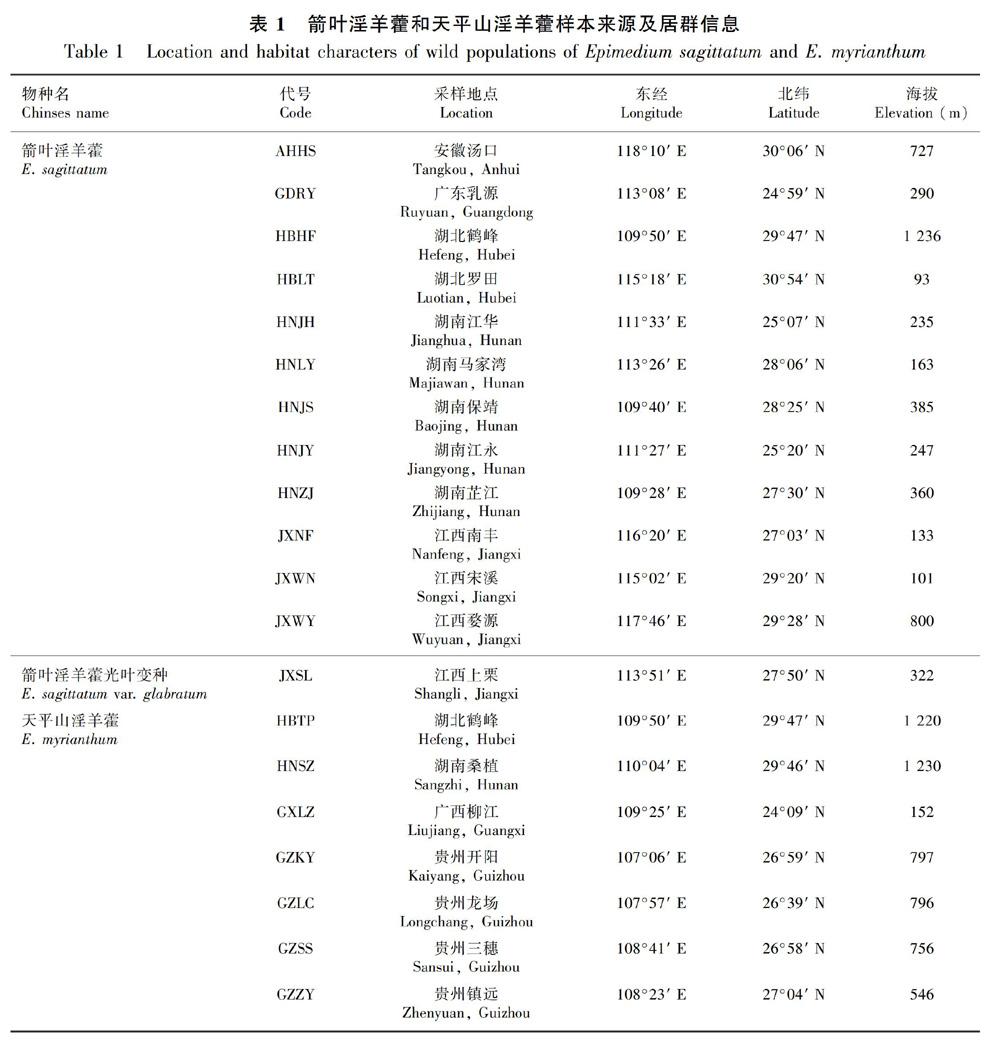
摘 要:该研究采用扫描电子显微镜(SEM,scanning electron microscope)对箭叶淫羊藿13个居群(含箭叶淫羊藿光叶变种1个居群)、天平山淫羊藿7个居群进行花粉形态和外壁纹饰的观察及对比研究。结果表明:两个类群的花粉均为近球形,具三孔沟,大小相近。箭叶淫羊藿的花粉大小为(18.21~21.66) μm × (19.55~23.63) μm,天平山淫羊藿的花粉大小为(18.32~21.43) μm × (19.73~23.48) μm。外壁纹饰是花粉形态的重要性状。不同居群间花粉外壁纹饰存在较大差异,主要分为网纹纹饰、条网纹纹饰和颗粒状纹饰三大类,继而根据网脊类型和网眼密度分为六亚类。其中,箭叶淫羊藿的花粉形态包含了全部的三大类六亚类,天平山淫羊藿包含其中的三大类四亚类。天平山淫羊藿花粉特征为箭叶淫羊藿花粉形态变异的子集。整体而言,外壁纹饰类型随纬度和海拔增加呈现出由网状纹饰向条网状纹饰过渡的趋势。花粉主要形态性状特征均与海拔存在密切相关性。在花粉形态及外壁纹饰特征上,天平山淫羊藿在箭叶淫羊藿变异范围之内,与其他宏观形态性状变异一致。花粉形态性状无法将两类群有效区分,未能提供天平山淫羊藿作為独立种的有力支撑。这暗示花粉形态证据在厘清近缘类群之间的系统发育关系意义相对有限,但能否对该属系统发育树大支的界定提供有力证据还有待于进一步研究。
关键词:居群,箭叶淫羊藿,天平山淫羊藿,花粉形态,形态变异
中图分类号:Q944
文献标识码:A
文章编号:1000-3142(2020)09-1300-15
Abstract:Epimedium myrianthum having much in common with E. sagittatum. There are transitions in many morphological characters and even in key taxonomy features. In order to explore the taxonomic relationship between the two taxa,the pollen morphology and ornamentation of exine of 13 populations of E. sagittatum (including 1 population of E. sagittatum var. glabratum T. S. Ying) and 7 populations of E. myrianthum were observed and compared by scanning electron microscope (SEM). The results showed that the pollen of both taxa was subspheroidal,with three-hole groove. Pollen size of E. sagittatum and E. myrianthum was (18.21-21.66) μm × (19.55-23.63) μm and (18.32-21.43) μm × (19.73-23.48) μm,respectively. The ornamentation of exine was an important character of pollen morphology. There were significant differences in ornamentation of exine among populations,which have been divided into three categories,namely reticulate,article reticulate and granular,and then divided into six subcategories according to the type of mesh ridge and mesh density. Among them,pollen morphology of E. sagittatum (including E. sagittatum var. glabratum) contained all three categories and six subcategories,and that of E. myrianthum contained three categories and four subcategories. This means that the pollen morphological variation pattern of E. myrianthum was a subset of E. sagittatum. On the whole,the variation pattern of the ornamentation of exine showed a trend,from reticulate to article reticulate,with the increase of latitude. And the main morphological characters of pollen were closely related to the elevation. As far as the pollen morphology concerned,the variation pattern of E. myrianthum was within the variation range of E. sagittatum,which was consistent with the variation patterns of other morphological characters. Pollen morphological characters could not distinguish the two taxa effectively,which failed to provide strong support for E. myrianthum as an independent species. It is suggested that the pollen morphological evidence is of relatively limited in clarifying the phylogenetic relationship among related taxa. However,further studies are still needed to provide strong evidence for the definition of large branches of phylogenetic trees of this genus.
Key words:population,E. sagittatum,E. myrianthum,pollen morphology,morphological variation
淫羊藿为小檗科(Berberidaceae)多年生草本植物,是我国传统且特有的重要药用植物,具有补肾阳、强筋骨和祛风湿等功效(国家药典委员会,2015),在改善心血管疾病、提高免疫力、防癌抗癌和抗抑郁等方面具有突出功效(Zhao et al.,2018; Choi et al.,2019)。淫羊藿属(Epimedium L.)全世界约68种,我国共发表58种,经修订后为46种,约85%的类群特有分布于中国(Xu et al.,2019; 徐艳琴等,2020)。淫羊藿属是分类上十分特殊和复杂的属,超过一半的物种为国外研究者根据少量栽培个体发表,1990年—1999年间,新种数量迅速增加,共发表26种且全部来自中国。由于短时间内物种数量的迅速增加,缺乏足够形态调查,加之种间杂交可育导致物种之间关系错综复雜,该属形成了大量复合种(species complex/species aggregate)(徐艳琴等,2014a)。其中,箭叶淫羊藿复合体(E. sagittatum species complex)为淫羊藿属最复杂的关注点之一,至少有10个类群与箭叶淫羊藿(E. sagittatum)关系密切(徐艳琴等,2014a)。徐艳琴等(2014b)对箭叶淫羊藿复合体的分类问题进行初步梳理,将毡毛淫羊藿(E. coactum)、龙头虎毡毛淫羊藿(E. coactum var. longtouhum)、裂叶淫羊藿(E. lobophyllum)、多花淫羊藿(E. multiflorum)、靖州淫羊藿(E. jingzhouense)、剑河淫羊藿(E. myrianthum var. jianaheense)处理为天平山淫羊藿(E. myrianthum)异名,贵州淫羊藿(E. sagittatum var. guizhouense)作为箭叶淫羊藿(E. sagittatum)异名,普定淫羊藿(E. pudingense)降为箭叶淫羊藿光叶变种(E. sagittatum var. glabratum),箭叶淫羊藿复合体的分类最终追溯到箭叶淫羊藿和天平山淫羊藿的关系。
箭叶淫羊藿是淫羊藿属分布范围较广的物种,也是形态变异最大、药用历史最悠久的物种,其分类学问题对淫羊藿属系统发育重建以及分类学研究具有重要意义(Stearn,2002; 郭宝林和肖培根,2003;徐艳琴,2008;徐艳琴等,2013,2014b)。此外,箭叶淫羊藿在历版《中华人民共和国药典》(以下简称药典)均有收录(国家药典委员会,2015),而天平山淫羊藿为非药典种,明确二者之间的分类关系对物种的准确鉴定、药用资源利用、药材质量和药效保障具有重要意义。不同学者从植物形态、遗传多样性、叶背非腺毛性状、化学活性成分和化学型等不同角度对箭叶淫羊藿进行分类学研究,结果显示其种内居群和个体间均呈现出丰富的变异(梁琼等,2013;徐艳琴等,2013;许瑛等,2015a,b)。
从20世纪90年代开始,陆续有学者对淫羊藿属植物花粉形态进行研究,如王悦云等(2015)对淫羊藿属31个物种花粉形态研究表明,本属植物的花粉大小和形状,尤其是外壁纹饰(网纹纹饰,条网纹纹饰和条纹状纹饰),可作为植物分类和药材鉴定的重要依据。但不同学者对箭叶淫羊藿花粉形态描述差异较大。余旭升和刘启宏(1991)描述的箭叶淫羊藿花粉粒径较大,为(29.7~32.6) μm × (32.4~37.5) μm,外壁具网状雕纹。梁海锐和阎文玫(1991)描述箭叶淫羊藿花粉大小(25~30) μm × (28.7~31.9) μm,外壁纹饰为条网状纹饰,网眼不规则。郭宝林等(1998)以及何顺志和郭宝林(1997)描述箭叶淫羊藿花粉大小为20.5 μm × (22.1~25.2) μm,外壁网状纹饰。三个产地间的箭叶淫羊藿花粉形态差异较大,花粉形态为长球形到近球形变化,外壁纹饰存在网脊形态和网眼密度的差异。王悦云等(2015)描述箭叶淫羊藿花粉大小(18.2~21.8) μm × (39.1~42.7) μm,外壁纹饰为网纹纹饰,网眼类圆形。
“居群”概念在分类学上至关重要,前期研究往往将少量样本代表一个物种,忽略了“居群”概念,无法获知种内居群间和个体间的差异。取样局限性导致对种间和种内变异范围和变异样式缺乏全面整体的把握,对性状的分类学价值难以正确判断,对“专属性”判断易存在偏差(徐艳琴等,2013,2014b;Xu et al.,2019),且导致不同学者研究结果很不一致。因此,本研究以“居群”为单位,对箭叶淫羊藿及其近缘种天平山淫羊藿展开系统的花粉形态研究,从花粉形态学角度为其分类关系提供参考。
1 材料与方法
1.1 材料
研究材料于花期严格按照居群采样,每个居群采集19~30株个体,经江西中医药大学徐艳琴教授鉴定,并于江西中医药大学进行同质园栽培。共采集20个居群,其中箭叶淫羊藿13个居群(含箭叶淫羊藿光叶变种1个居群),天平山淫羊藿7个居群。居群覆盖物种主要分布区,包括湖南、湖北、贵州、江西、安徽、广东和广西等7省(区)。居群采集位置、代号、经纬度和海拔高度等信息见表1。花粉观测于2019年花期(3月—4月)取自同质园栽培植株。每个居群随机选取5~6株个体,采集10~20朵雄蕊成熟的花,浸泡于FAA溶液(38%甲醛5 mL,冰醋酸5 mL,70%酒精90 mL)中,避光保存,备用。
1.2 方法
取少量花粉使用50%~90%浓度的乙醇梯度洗脱,干燥后用牙签挑取少量花粉,均匀粘在样品台的导电胶纸上,置于FEI型Quanta 250场发射环境扫描电子显微镜(SEM)下进行观察,工作电压为5 kV。在不同倍数下选取清晰且具有代表性的视野进行观测,于800倍视野下观察花粉整体效果并随机选取20粒花粉测量其极轴长(polar axis)和赤道轴长(equator axis),10 000倍视野下观察花粉的极面和赤道面,24 000倍视野下观察花粉外壁纹饰特征,并测量网眼直径(mesh diameter)和网脊宽度(mesh ridge width)。不同视野均拍照保存。
1.3 数据统计分析
计算极轴长与赤道轴长的比值(P/E,Polar axis/equator axis),网眼直径与网脊宽度比值(D/W,Diameter/width)。用SPSS21.0软件进行单因素方差分析,统计各居群花粉形态性状的平均值及标准差,将三种花粉外壁纹饰(网纹纹饰、条网纹纹饰、颗粒状纹饰)分别赋值为1、2、3,对花粉外壁纹饰和主要地理生态因子(经度、纬度与海拔)进行相关性分析。用MVSP3.0软件对花粉外壁类型及形态量化性状进行数据标准化及聚类分析。用DPS9.01软件进行灰色关联度分析,花粉形态特征作为母因素,生态因子作为子因素。用Photoshop软件编辑图片。为保证研究的系统性和可比性,本研究花粉形态特征的描述术语及类型划分参考梁海锐和阎文玫(1991)、郭宝林等(1998)和王悦云等(2015)对淫羊藿属植物花粉形态的研究。
2 结果与分析
2.1 箭叶淫羊藿与天平山淫羊藿花粉形态比较 (图5)
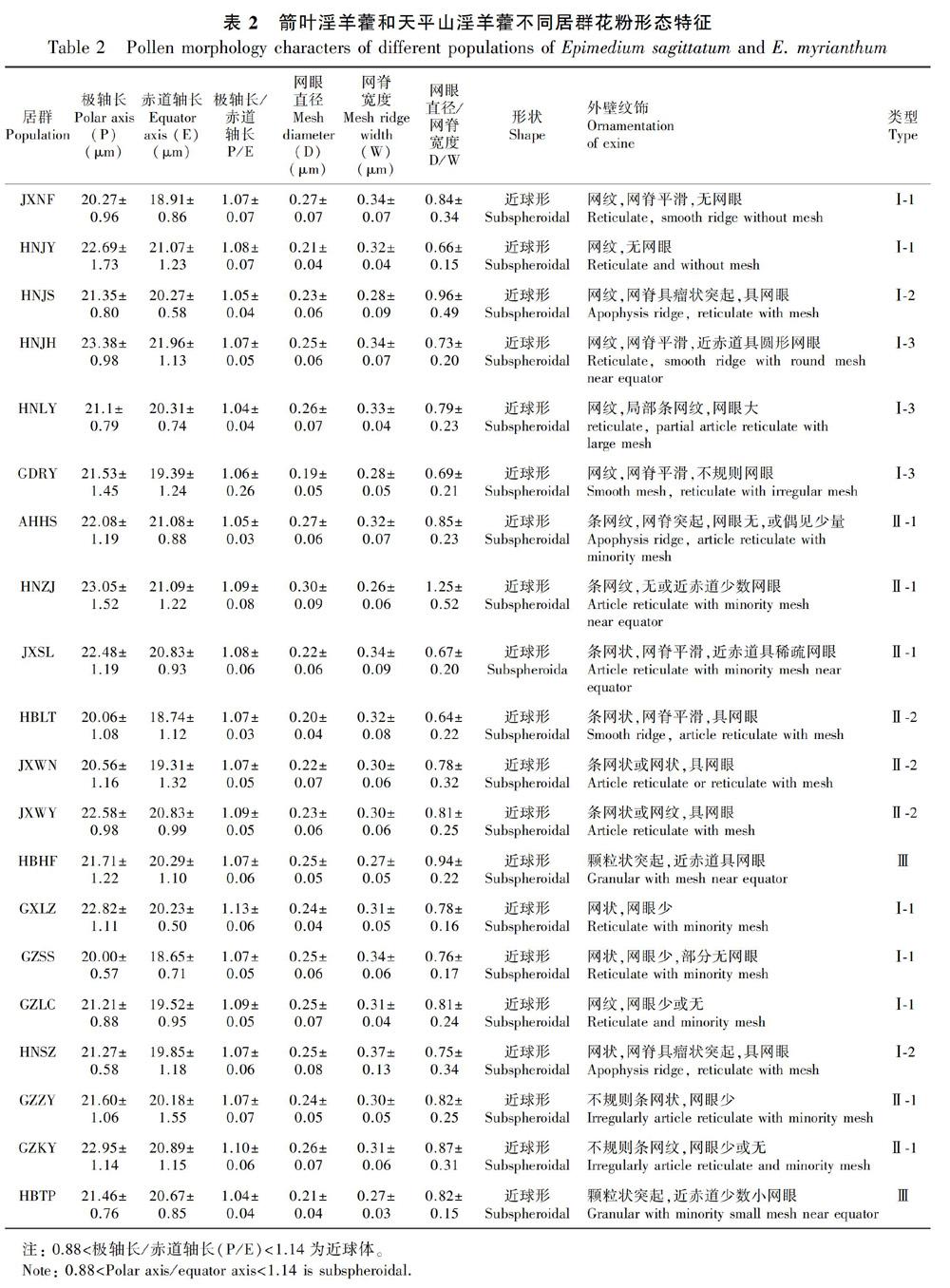
2.1.1花粉大小 箭叶淫羊藿和天平山淫羊藿花粉均为单粒,近球形,具三裂孔沟。两种淫羊藿花粉大小差异不大,箭叶淫羊藿花粉粒大小为 (18.21~21.66) μm × (19.55~23.63) μm,P/E为1.06~1.08,天平山淫羊藿花粉粒大小为(18.32~21.43) μm × (19.73~23.48) μm,P/E为1.07~1.09。
2.1.2 花粉外壁纹饰 根据花粉外壁纹饰可以分为三大类:网状纹饰(Ⅰ)、条网状纹饰(Ⅱ)和颗粒状突起纹饰(Ⅲ)。
第Ⅰ类为网纹纹饰。根据网脊和网眼密度,该类型进而分为三个亚类:Ⅰ-1类,网纹纹饰,网脊平滑,无网眼或具稀疏网眼,此类型包括箭叶淫羊藿JXNF和HNJY居群,天平山淫羊藿GXLZ、GZSS和GZLC居群;Ⅰ-2类,网纹纹饰,网脊具瘤状突起,具网眼,此类型包括箭叶淫羊藿HNJS居群和天平山淫羊藿HNSZ居群;Ⅰ-3类,网纹纹饰,网脊平滑,网眼多,此类型包括箭叶淫羊藿HNJH、HNLY和GDRY居群,其中GDRY和HNLY两个居群花粉外壁纹饰均出现局部不规则条网状纹饰。
第Ⅱ类为条网纹纹饰。该类型根據纹饰方向和网眼密度进而分为两个亚类:Ⅱ-1类,方向不规则条网纹纹饰,网脊平滑,网眼较少或无,包括箭叶淫羊藿AHHS和HNZJ居群、箭叶淫羊藿光叶变种JXSL居群、天平山淫羊藿GZZY和GZKY居群;Ⅱ-2类,方向规则条网状纹饰,网脊平滑,网眼较多,包括箭叶淫羊藿HBLT、JXWN和JXWY居群。其中:JXWN居群1/3的花粉外壁纹饰为条网纹纹饰,2/3的花粉其外壁纹饰以条网纹纹饰为主,局部呈现网状纹饰;JXWY居群花粉外壁纹饰为条网状纹饰,局部呈现网状纹饰。
第Ⅲ类为颗粒状纹饰。花粉外壁具颗粒状突起纹饰,无网眼,包括箭叶淫羊藿HBHF居群和天平山淫羊藿HBTP居群。
由以上结果可知,箭叶淫羊藿和天平山淫羊藿两类群的花粉形态均存在较丰富的变异,均包含了三类外壁纹饰。但是,箭叶淫羊藿出现了所有六亚类类型,天平山淫羊藿则包含其中四亚类。因此,天平山淫羊藿的花粉形态性状为箭叶淫羊藿花粉形态变异的子集。
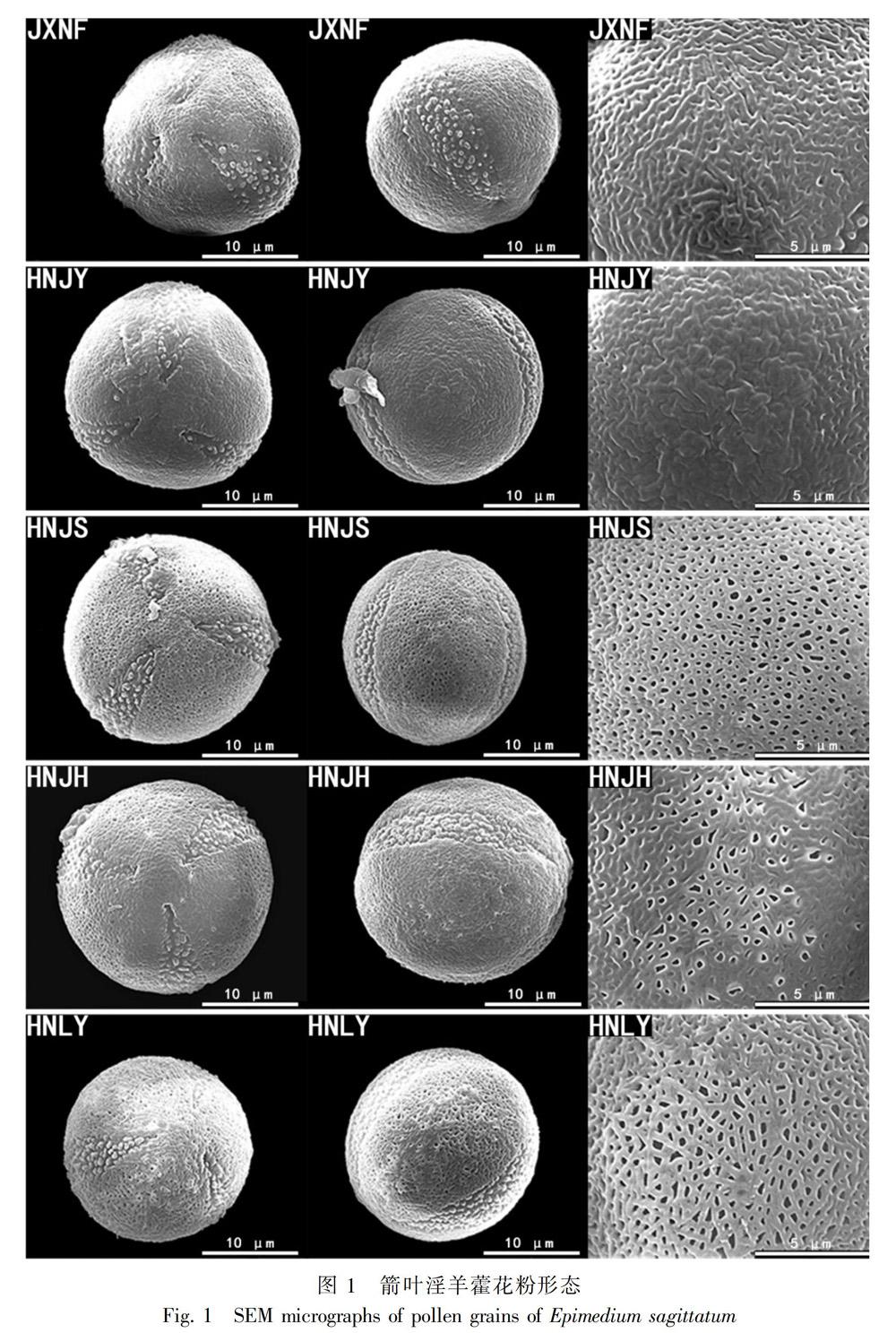
2.2 箭叶淫羊藿不同居群的花粉形态 (图1,图2,图3)
2.2.1 网纹纹饰 (Ⅰ类)
2.2.1.1 网纹纹饰-网脊平滑,网眼无或稀疏(Ⅰ-1类) JXNF花粉粒大小(18.51~19.32) μm × (19.82~20.72) μm,近球形或近扁球形,两极平坦,极面观呈三裂,近三角圆形,赤道面呈近圆形。沟膜具稀疏颗粒状突起,突起顶端圆钝,沟较宽。花粉外壁凹凸不平,网纹纹饰,网脊凸起且光滑,无网眼,近两极与沟边处网纹模糊。HNJY 花粉粒大小(20.50~21.64) μm × (21.88~23.50) μm,近球形,两极平坦,极面观三裂,圆形,赤道面呈近圆形。沟膜具瘤状突起,分布均匀。花粉外壁凹凸不平,网纹纹饰,网纹较模糊,无网眼,网脊表面无突起。
2.2.1.2 网纹纹饰-网脊具瘤状突起,网眼较多(Ⅰ-2类) HNJS 花粉粒大小(20.00~20.54) μm × (20.97~21.72) μm,近球形,极面观圆形,具三沟,中间较宽。沟膜褶皱,具颗粒状突起。外壁具清晰网状纹饰,网脊上表面不平,具瘤状突起,网眼密集,类圆形,分布均匀,近两极网眼变小,网脊宽度与网眼直径相近。
2.2.1.3 网纹纹饰-网脊平滑,网眼较多(Ⅰ-3类) HNJH 花粉粒大小(21.69~22.64) μm × (23.09~24.07) μm,近球形,极面观三裂圆形,极面观较大,赤道面呈椭圆形。沟较宽,沟膜具颗粒状突起,分布均匀,颗粒顶端圆钝。花粉外壁网状纹饰,近赤道轴具大小不一的网眼。但靠近两极和沟边无网眼,外壁部分区域也出现模糊无网眼的纹饰。HNLY 花粉粒大小(19.97~20.66) μm × (20.73~21.47) μm,近球形,极面观三裂圆形,赤道面两极平坦。沟膜具颗粒状突起,顶端圆钝,分布均匀。花粉外壁网状纹饰局部呈条网状纹饰,网纹清晰,近赤道轴网眼较大,不规则均匀分布,偶可通过网眼看见柱状层。近两极与两沟边处网眼较小。
GDRY 花粉粒大小(18.79~19.98) μm × (20.83~22.23) μm,近球形,极面观三裂圆形,沟膜具较大颗粒状突起。花粉外壁网状纹饰局部呈不规则条网状纹饰,具大小不一形状不规则的网眼。需特别说明的是,GDRY和HNLY两个居群出现了花粉纹饰的变异,主要为网状纹饰但局部呈现不规则条网状纹饰。根据外壁纹饰整体情况,将其仍划分到网纹纹饰类型下。
2.2.2 条网纹纹饰 (Ⅱ类)
2.2.2.1条网纹纹饰-网眼少或无,不规则条网纹(Ⅱ-1类) AHHS花粉粒大小(20.67~21.49) μm × (21.53~22.64) μm,近球形,极面观三裂圆形,赤道面椭圆形。沟膜密布均匀颗粒状突起,顶端圆钝。花粉外壁条网状纹饰,网纹方向不规则,网纹清晰,方向不规则,条状网脊突起,两极与两沟边条网状纹饰模糊,外壁偶见极少数不均匀小网眼。HNZJ 花粉粒大小(20.52~21.66) μm × (22.33~23.76) μm,极面观三裂圆形,沟膜具密集瘤状突起。花粉外壁为条网状纹饰,网纹方向不规则,无网眼或近赤道轴处具极稀疏网眼。JXSL花粉粒大小(20.39~21.26) μm × (21.92~23.04) μm,近球形,极面观三裂圆形,赤道面两极渐窄。沟较窄,沟膜具颗粒状突起,顶端圆钝,分布均匀。花粉外壁条网状纹饰,网纹清晰,网脊平滑,赤道轴稀疏网眼,分布不均匀,近两极与两沟边无网眼。
2.2.2.2 条网纹纹饰-网眼多 (Ⅱ-2类)HBLT 花粉粒大小(18.22~19.27) μm × (19.55~20.56)μm,近球形或长球形,极面观圆形,赤道面两端渐窄,中间较宽。沟膜颗粒密集,分布均匀,颗粒顶端圆钝。外壁纹饰条网状纹饰,网纹沿极轴方向延伸,网脊平滑密集。网眼较小,不规则,网脊宽度大于网眼直径,近赤道轴网眼多,近沟与两极部位外壁网眼变小、变少。JXWN 花粉粒大小(18.69~19.93) μm × (20.01~21.10) μm,近球形,极面观三裂圆形,两极平坦。沟较窄,沟膜具颗粒状突起,顶端圆钝。花粉外壁条网状纹饰或网状纹饰,网纹清晰,方向不规则,网眼密集,分布均匀,近两极或沟边网眼变小。部分花粉粒近赤道轴无网眼,在近两沟具稀疏网眼。JXWY花粉粒大小(20.51~21.14) μm × (22.27~22.90) μm,近球形,极面观三裂圆形,两极平坦。沟较窄,沟膜具颗粒状突起,顶端圆钝。花粉外壁条网状纹饰,局部网状纹饰,网纹清晰,近赤道轴具大小不一、形状不规则网眼,近两极或沟边无网眼。
需特别说明的是,JXWN和JXWY两个居群出现了花粉纹饰的变异,JXWN居群1/3的花粉外壁纹饰为条网纹纹饰,2/3的花粉其外壁纹饰以条网纹纹饰为主,局部呈现网状纹饰。JXWY居群花粉外壁纹饰为条网状纹饰,局部呈现网状纹饰。
2.2.3 颗粒状突起 (Ⅲ类) HBHF 花粉粒大小(19.77~20.81) μm × (21.14~22.28) μm,近球形,极面观三裂圆形,赤道面呈近圆形。沟膜具不规则颗粒状突起。花粉外壁凹凸不平,密布瘤状小突起,两极具极稀疏网眼,部分近赤道轴具极少数几个网眼,网眼极小,网脊光滑。
2.3 天平山淫羊藿花粉形态 (图3,图4)
2.3.1 网纹纹饰(Ⅰ类)
2.3.1.1 网纹纹饰-网眼少或无 (Ⅰ-1类) GXLZ 花粉粒大小(19.99~20.46) μm × (22.30~23.34) μm,近球形,極面观三裂圆形,赤道面两极渐窄。沟较窄,近两极渐尖或极尖,沟膜具密集不规则颗粒状突起,顶端圆钝,分布均匀。花粉外壁网状纹饰,网纹清晰,具极稀疏小网眼。GZSS 花粉粒大小(18.32~18.98) μm × (19.73~20.27) μm,近球形,极面观三裂圆形,赤道面两极渐窄。沟较窄,近两
极渐尖或极尖,沟膜具密集不规则颗粒状突起。花粉外壁网状纹饰,局部具稀疏大小不一形态不规则的网眼。部分花粉粒外壁网纹模糊,无网眼。GZLC 花粉粒大小(19.08~19.97) μm × (20.80~21.63) μm,近球形,极面观三裂圆形。沟较宽,沟膜具颗粒状突起。花粉外壁网状纹饰,部分花粉粒网纹模糊。无网眼或近赤道轴具稀疏不规则网眼。
2.3.1.2 网纹纹饰-网眼较多,网脊具瘤状突起(Ⅰ-2类)HNSZ 花粉粒大小(19.10~20.60) μm × (20.90~21.64) μm,近球形,极面观三裂圆形。
沟膜具颗粒状突起。花粉外壁网状纹饰,网纹清晰,网脊上具密集瘤状突起,赤道轴具大小不一、近圆形的网眼。
2.3.2 条网纹纹饰 (Ⅱ类)
条网纹纹饰-网眼少或无 (Ⅱ-1类) GZZY 花粉粒大小(19.46~20.90) μm × (21.10~22.09) μm,近球形,极面观三裂圆形,赤道面近两极渐窄。沟膜具颗粒状突起。花粉外壁方向不规则条网状纹饰,网纹清晰,近赤道轴具极稀疏不均匀小网眼。GZKY 花粉粒大小(20.35~21.43) μm × (22.41~23.48) μm,近球形,极面观三裂圆形,赤道面两极渐窄。沟较窄,近两极渐尖或极尖,沟膜具密集不规则颗粒状突起。花粉外壁具不规则条网状纹饰,近两极与沟边网纹模糊,偶见具极稀疏小网眼。
2.3.3颗粒状突起 (Ⅲ类) HBTP 花粉粒大小(20.27~21.06) μm × (21.10~21.81) μm,近球形,极面观三裂圆形。沟较窄,近两极渐尖,沟膜具密集不规则颗粒状突起。花粉外壁具密集颗粒状突起,赤道轴具极稀疏不均匀小网眼。
2.4 花粉形态特征与生态因子相关性
2.4.1 花粉外壁纹饰与生态因子的相关性分析 从各居群的地理分布及其花粉外壁纹饰特征来看,两者呈现一定规律(图5)。网状纹饰(Ⅰ型)主要分布于广东和广西的北部,贵州、江西和湖南三省南部,如GZLZ、HNJY、HNJH、GZLC、GZSS和JXNF居群。条网状纹饰(Ⅱ型)主要分布于贵州、湖南和江西三省的中部,安徽省南部及邻近的湖北省区域,如GZKY、GZZY、HNZJ、JXSL、AHHS和HBLT居群。颗粒状纹饰(Ⅲ型)的HBHF和HBTP两个居群分布于湖北西部。从整体来看,花粉外壁纹饰呈现随纬度逐渐过渡的趋势。与海拔、经度及纬度的相关性分析结果显示,纬度与花粉外壁纹饰类型显著相关(P=0.006),随着纬度升高,花粉外壁纹饰从Ⅰ型(网状纹饰)过渡到Ⅱ型(条网状纹饰)型和Ⅲ型(颗粒状纹饰)。花粉外壁纹饰类型与海拔呈显著相关(P=0.046),与经度不存在显著相关性(P=0.829)。
2.4.2 花粉形态特征与生态因子的灰色关联度分析 对不同居群淫羊藿花粉形态特征性状与生态因子灰色关联度分析结果表明,不同生态因子对花粉形态性状的影响各不同(表3)。其中,对赤道轴长、网孔直径、网眼直径/网脊宽度和花粉外壁纹饰影响程度均为海拔>北纬>东经,对极轴长和极轴长/赤道轴长影响程度为海拔>东经>北纬,对网脊宽度影响程度为东经>海拔>北纬。
三角形:箭叶淫羊藿(含光叶变种); 正方形:天平山淫羊藿。绿色:网状纹饰; 红色:条网状纹饰; 黑色:颗粒状纹饰; 红色为主加绿色(JXWN、JXWY):主要为条网纹纹饰,局部出现网纹纹饰; 绿色为主加红色(GDRY、HNLY):主要为网纹纹饰,局部出现条网纹纹饰。实心:网眼少或无; 空心:网眼较多。下同。
triangle:Epimedium sagittatum and E. sagittatum var. glabratum; square:E. myrianthum. green:Reticulate exine; red:Article reticulate exine; black:Granular exine; red plus green (JXWN,JXWY):Mainly article reticulate exine,with reticulate exine; green plus red (GDRY,HNLY):Mainly reticulate exine,with article reticulate exine. solid:Few or no pollen mesh; hollow:With many pollen meshes. The same below.
从总体来看,淫羊藿花粉形态特征指标与生态因子之间的灰色关联系数(ri)为0.589 6~0.734 6,表明生态因子与淫羊藿花粉形态关系较为密切。其中,海拔与极轴长、赤道轴长、极轴长/赤道轴长、网眼直径、网眼直径/网脊宽度和花粉纹饰类型的ri均为最高,说明海拔是影响花粉主要形态性状的主导因子(ri= 0.665 0~0.734 6);东经是影响网脊宽度的主导因子(ri= 0.638 8)。值得关注的是,北纬是影响花粉形态大小特征和花粉外壁纹饰的重要因子(ri = 0.589 6~0.712 0)。
基于以上花粉形态特征与生态因子相关性分析结果表明,两者间存在密切相关性。就外壁纹饰及生态因子相关性而言,不同分析软件结果存在一定差异。SPSS21.0分析结果显示,纬度和海拔均与花粉外壁纹饰存在显著相关性,但纬度的相关性更强。DPS9.01分析结果显示,海拔和纬度均与花粉外壁纹饰关系密切,但海拔的关联度更高。就整体而言,以上均说明海拔和纬度对花粉形态的影响较突出。
2.5 花粉形态聚类
聚类分析结果显示,在遗传距离3.2处,所有居群分为五组(图6)。由图6可知,HNZJ花粉粒较大,单独聚为一组;第二组包括箭叶淫羊藿HBHF,天平山淫羊藿HBTP,花粉外壁纹饰类型为颗粒状纹饰;第三组包括箭叶淫羊藿4个居群(HNJH、JXSL、HNJY、JXWY)和天平山淫羊藿2个居群(GXLZ、GZKY),包含了网状纹饰和条网状纹饰类型;第四组为箭叶淫羊藿HBLT,花粉外壁纹饰为条网状纹饰;第五组包括箭叶淫羊藿6个居群(JXNF、GDRY、HNJS、JXWN、HNLY和AHHS),天平山淫羊藿4个居群(HNSZ、GZSS、GZLC和GZZY),花粉外壁纹饰包括网纹纹饰和条网纹纹饰类型。
基于花粉形态数据的聚类分析结果表明,箭叶淫羊藿与天平山淫羊藿在花粉形态特征上无明显区分和间断,两类群的居群在聚类分支中混杂。3 讨论与结论
3.1 箭叶淫羊藿花粉形态特征及分类启示
箭叶淫羊藿作为淫羊藿属植物中种内变异范围最大的物种,无论是在植物形态、遗传变异,还是在化学型上都存在丰富的多样性,可能是多型种(徐艳琴,2008;梁琼等,2013;许瑛等,2015a)。暗示对箭叶淫羊藿的研究,样本的选择需非常谨慎(徐艳琴等,2013)。样本要尽可能全面,在样本数量不足的情况下,一个连续的变异式样会呈现间断的形式,种及种下类群的建立应以天然居群为基础,这是现代分类学的基本原则(汤彦承和向秋云,1989)。因为同一物种的居群间甚至同一居群不同个体间都可能存在一定程度变异,以居群为基础的系统研究对全面掌握类群变异范围和变异式样,评估其分类价值具有重要意义(刘少雄等,2016;Liu et al.,2017; Xu et al.,2019)。但是,以往淫羊藿属孢粉学研究往往忽略“居群”概念,以单一产地的少量代表整个物种,导致不同学者对同一物种花粉形态描述存在较大差异,且在分类上易出现分类过细的弊端。
本研究对箭叶淫羊藿12个居群及其变种1居群的花粉形态进行系统研究,发现花粉性状与宏观形态变异相似,同样存在广泛变异,不同居群在其主要特征上存在较大差异。观察到网状纹饰、条网状纹饰和颗粒状纹饰三大类花粉外壁纹饰,进而根据网脊和网眼特征细分为六亚类。从花粉外壁纹饰性状来看,本研究涵盖了以往研究的所有变异,如梁海锐和阎文玫(1991)观察的条网状纹饰,余旭升和刘启宏(1991)报道的网状雕纹,何顺志和郭宝林(1997)、王悦云等(2015)描述的网状纹饰。此外,本研究还观察到条网状纹饰和颗粒状纹饰。这证明箭叶淫羊藿研究取材全面的重要性和必要性。
王悦云等(2015)基于淫羊藿属31种植物花粉形态的研究,认为花粉外壁纹饰可用于植物分类的依据,并根据箭叶淫羊藿花粉外壁为网纹纹饰,而普定淫羊藿花粉外壁为条网纹纹饰,从而支持普定淫羊藿独立成种。但从本研究结果来看,箭叶淫羊藿多个居群花粉外壁纹饰都为条网状纹饰(如AHHS、HNZJ、HBLT、JXWN和JXWY),证明该性状并非为普定淫羊藿特有的间断性状,因此依据花粉特征不能支持普定淫羊藿单独成种。徐艳琴等(2014b)根据宏观外部形态特征,将普定淫羊藿降为箭叶淫羊藿光叶变种。王悦云等(2015)描述的普定淫羊藿与本研究中箭叶淫羊藿光叶变种JXSL居群的花粉外壁形态均为条网状纹饰,依据花粉形态更支持将普定淫羊藿降为箭叶淫羊藿光叶变种。
3.2 箭叶淫羊藿和天平山淫羊藿分类关系讨论
箭叶淫羊藿复合体的分类问题最终追溯到箭叶淫羊藿和天平山淫羊藿之间的关系上(徐艳琴等,2014b)。本课题组基于广泛的野外形态观测,发现两类群间多个关键性状重叠,如Stearn(1998)描述天平山淫羊藿新种时指出,箭叶淫羊藿叶缘波状,幼时绿色,而天平山淫羊藿叶缘平展,幼时带有明显红色斑点。但实际两类群多数居群幼叶均带有数量和深浅不一红色斑点或斑块。此外,在花茎上叶的数量、外萼颜色、花瓣颜色和形状等方面,两类群均存在过渡和交集(徐艳琴等,2014b)。两类群最主要的区别在于花序类型,是狭而直的圆锥花序(箭叶淫羊藿)还是铺伞形圆锥花序(天平山淫羊藿),以及由此导致小花数量和大小上的差异(徐艳琴等,2014b)。许瑛等(2015a,b,2016)先后基于淫羊藿多苷成分淫对箭叶淫羊藿的16个居群和天平山淫羊藿的8个居群进行质量评价,发现两类群不同居群间均存在较大差异,且天平山淫羊藿三种化学型在箭叶淫羊藿化学型变异范围内。本研究基于花粉形态数据及聚类分析结果显示,天平山淫羊藿与箭叶淫羊藿在花粉形态特征上无明显区分和间断,外壁纹饰的变异也均在箭叶淫羊藿变异范围之内,与其宏观形态性状变异趋势一致。因此,基于孢粉学证据,天平山淫羊藿与箭叶淫羊藿不存在明显间断,未能提供天平山淫羊藿单独成种的有力证据。但目前尚不能因为天平山淫羊藿的花粉形态变异在箭叶淫羊藿变异范围内,就否定天平山淫羊藿的物种地位。从前人对淫羊藿属不同物种花粉形态的研究结果来看(梁海锐和阎文玫,1991;余旭升和刘启宏,1991;何顺志和郭宝林,1997;郭宝林等,1998;王悦云等,2015),本研究箭葉淫羊藿花粉外壁纹饰的变异涵盖了多数物种的变异。因此,孢粉学证据的分类学意义在于对整属的花粉形态特征只有进行系统研究,才能获得客观结论。
参考文献:
CHOI H,KIM J,LI Z,et al.,2019. Effectiveness of prenyl group on flavonoids from Epimedium koreanum Nakai on bacterial neuraminidase inhibition [J]. Molecules,24(2):317.
GUO BL,XIAO PG,2003. Comment on main species of herba Epimedii [J]. Chin J Chin Mat Med,28(4):303-339. [郭宝林,肖培根,2003. 中药淫羊藿主要品类评述 [J]. 中国中药杂志,28(4):303-307.]
GUO BL,XIAO PG,HE SZ,1998. The pollen exine ornamentation of Epimedium and its taxonomic significance [J]. Acta Bot Yunnan,20(2):220-224. [郭宝林,肖培根,何顺志,1998. 花粉的外壁纹饰及分类学意义 [J]. 云南植物研究,20(2):220-224.]
HE SZ,GUO BL,1997. Scanning electron microscopy of pollen morphology on Epimedium medicinal plant in Guizhou [J]. Chin J Chin Mat Med,22(5):268-270. [何顺志,郭宝林,1997. 贵州产淫羊藿属药用植物花粉形态的扫描电镜观察 [J].中国中药杂志,22(5):268-270.]
LIANG HR,YAN WM,1991. Study on the pollen morphology of Epimedium in China [J]. Bull Bot Res,11(4):81-92. [梁海锐,阎文玫,1991. 国产淫羊藿属花粉形态研究 [J]. 植物研究,11(4):81-92.]
LIANG Q,ZHANG YJ,XU YQ,et al.,2013. Morphological variations and genetic diversity of Epimedium sagittatum populations [J]. Plant Sci J,31(4):422-427. [梁琼,张燕君,徐艳琴,等,2013. 箭叶淫羊藿居群形态及遗传多样性比较研究 [J]. 植物科学学报,31(4):422-427.]
LIU SX,LIU LJ,HUANG XF,et al.,2017. A taxonomic revision of three Chinese spurless species of genus Epimedium L. (Berberidaceae) [J]. PhytoKeys,78:23-36.
LIU SX,SHI HJ,XU YQ,2016. Morphological comparison of the Epimedium franchetii Stearn species complex based on population observation and implications for taxonomy [J]. Plant Sci J,34(3):325-339. [刘少雄,石慧君,徐艳琴,2016. 基于居群观测的木鱼坪淫羊藿复合种形态比较和分类学启示 [J]. 植物科学学报,34(3):325-339.]
National Pharmacopoeia Commission,2015. Pharmacopoeia of the Peoples Republic of China (Volume I) [S]. Beijing:China Medical Science Press:327. [國家药典委员会,2015. 中华人民共和国药典(一部) [S]. 北京:中国医药科技出版社:327.]
STEARN WT,1998. Four more Chinese species of Epimedium (Berberidaceae) [J]. Kew Bull,53(1):213-223.
STEARN WT,2002. The genus Epimedium and other herbaceous Berberidaceae,including the genus Podophyllum. A botanical magazine monograph [M]. Oregon:Timber press:26-351.
TANG YC,XIANG QY,1989. A reclassification of the genus Clematoclethra (Actinidiaceae) and further note on the methodology of plant taxonomy [J]. Acta Phytotax Sin,27(2): 81-95. [汤彦承,向秋云,1989. 重订藤山柳属的分类——续谈植物分类学工作方法 [J]. 植物分类学报,27(2):81-95.]
WANG YY,SUN QW,YANG W,et al.,2015. Pollen characteristics and taxonomic significance of Chinese Epimedium medicinal plant [J]. Acta Bot Boreal-Occident Sin,35(7):1367-1377. [王悦云,孙庆文,杨雯,等,2015. 中国淫羊藿属药用植物花粉特征及其分类学意义 [J]. 西北植物学报,35(7):1367-1377.]
XU CY,HOU YX,LI XH,et al.,2019. Pollen morphological variation of Primula merrilliana and its systematic significance [J]. Flora,253:43-48.
XU Y,SHI HJ,XU QY,2015a. ABCI chemical patterns of Epimedium sagittatum based on 16 wild populations:implication for utilization [J]. Lishizhen Med Mat Med Res,26(8): 2002-2006. [许瑛,石慧君,徐艳琴,2015a. 箭叶淫羊藿不同居群淫羊藿多苷的变异研究及利用启示 [J]. 时珍国医国药,26(8):2002-2006.]
XU Y,SHI HJ,LIU SX,et al.,2015b. Quality evaluation of Epimedium sagittatum in different populations based on icariin contents [J]. Chin Trad Herb Drugs,46(15):2284-2290. [许瑛,石慧君,刘少雄,等,2015b. 不同居群箭叶淫羊藿基于淫羊藿多苷成分的质量评价 [J]. 中草药,46(15):2284-2290.]
XU Y,SHI HJ,LIU SX,et al.,2016. Variation of ABCI in Epimedium myrianthum Stearn based on eight wild populations [J]. Lishizhen Med Mat Med Res,27(7):1720-1723. [许瑛,石慧君,刘少雄,等,2016. 天平山淫羊藿不同居群基于淫羊藿多苷成分的质量评价 [J]. 时珍国医国药,27(7):1720-1723.]
XU YQ,HUANG H,JIANG Y,et al.,2020. Taxonomic study of Epimedium L.:status,issues and prospect [J]. Guihaia,40(5):601-617. [徐艳琴,黄华,蒋勇,等,2020. 淫羊藿属分类学研究:进展、问题与展望 [J]. 广西植物,40(5):601-617.]
XU YQ,LIU LJ,LIU SX,et al.,2019. The taxonomic relevance of flower colour for Epimedium (Berberidaceae),with morphological and nomenclatural notes for five species from China [J]. PhytoKeys,118:33-64.
XU YQ,XU Y,LIU Y,et al.,2014a. Progress and open problems in classical taxonomic research on Epimedium L. [J]. Chin Trad Herb Drugs,45(4):569-577. [徐艷琴,许瑛,刘勇,等,2014a. 淫羊藿属经典分类学研究进展及存在的问题 [J]. 中草药,45(4):569-577.]
XU YQ,XU Y,SHI HJ,et al.,2014b. Taxonomic research on Epimedium sagittatum species complex and discussion [J]. Chin Trad Herb Drugs,45(22):3343-3349. [徐艳琴,许瑛,石慧君,等,2014b. 箭叶淫羊藿复合体的分类问题及讨论 [J]. 中草药,45(22):3343-3349.]
XU YQ,2008. Determination of active constituents and population genetics of three medicinal Epimedium species [D]. Wuhan:Wuhan Botanical Garden,Chinese Academy of Sciences. [徐艳琴,2008. 三种药用淫羊藿的活性成分变异及居群遗传学分析 [D]. 武汉:中国科学院武汉植物园.]
XU YQ,CAI WZ,HU SF,et al.,2013. Morphological variation of non-glandular hairs in cultivated Epimedium sagittatum (Berberidaceae) populations and implications for taxonomy [J]. Biodivers Sci,21(2):185-196. [徐艳琴,蔡婉珍,胡生福,等,2013. 箭叶淫羊藿同质园栽培居群非腺毛多样性及其分类学启示 [J]. 生物多样性,21(2):185-196.]
XU YQ,LI ZZ,ZHANG XJ,et al.,2008. Geographic distribution and resource status of three important medicinal Epimedium species [J]. J Wuhan Bot Res,26(1):91-98. [徐艳琴,李作洲,张学军,等,2008. 三种药用淫羊藿的地理分布与资源调查 [J]. 武汉植物研究,26(1):91-98.]
YU XS,LIU QH,1991. A study on the pollen morphology of Epimedium in Hubei Province [J]. J Wuhan Bot Res,9(3): 295-299. [余旭升,刘启宏,1991. 湖北淫羊藿属植物花粉形态研究 [J]. 武汉植物学研究,9(3):295-299.]
ZHAO Y,CHEN S,WANG Y,et al.,2018. Effect of drying processes on prenylflavonoid content and antioxidant activity of Epimedium koreanum Nakai [J]. J Food Drug Anal,26(2): 706-806.
(责任编辑 蒋巧媛)
猜你喜欢
植物研究(2022年5期)2022-10-12 06:40:10
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
西北植物学报(2021年2期)2021-03-29 07:54:52
中国粮油学报(2020年12期)2021-01-09 05:15:12
草地学报(2018年5期)2018-11-07 02:25:00
草地学报(2018年1期)2018-07-30 03:29:06
皖西学院学报(2018年2期)2018-05-21 11:25:45
山西农业大学学报(自然科学版)(2016年10期)2016-10-21 01:24:03
植物资源与环境学报(2012年1期)2012-12-31 13:31:26
