
斜纹夜蛾核多角体病毒Splt75基因的生物信息学分析
2020-10-30 05:24李赛男吕怡娜甘田刘文华
生物化工 2020年5期
李赛男,吕怡娜,甘田,刘文华
(肇庆学院生命科学学院,广东肇庆 526061)
杆状病毒是一类特异性感染节肢动物的病原微生物,其典型特征是具有双向复制周期,在复制周期中产生两种不同形态和功能的病毒粒子,分别为芽生型病毒粒子(Budded virion,BV)和包埋型病毒粒子(Occlusion-derived virion,ODV)。这两种病毒粒子的囊膜结构和组分不同,但是核酸相同以及核衣壳组成相近[1-3]。BV负责建立系统感染,ODV负责在昆虫宿主中的水平传播[3-8]。杆状病毒在应用上主要作为外源基因的高效表达载体和生物杀虫剂[9]。
斜纹夜蛾(Spodoptera litura)俗称夜盗虫、黑头虫,又名莲纹夜蛾,属鳞翅目夜蛾科,是一种杂食性农业害虫,危害的植物达到109科389种,在亚洲、非洲、欧洲、大洋洲等地均有分布,经常造成大范围的严重灾害[10]。目前,斜纹夜蛾的防治方式主要有生物防治、诱杀成虫和化学防治等。斜纹夜蛾核多角体病毒(Spodoptera litura multicapsid nucleopolyhedrovirus,SpltMNPV)能特异高效地控制斜纹夜蛾种群,已被中山大学有害生物控制与资源利用国家重点实验室开发成为商品化病毒杀虫剂[11]。杆状病毒作为生物杀虫剂,相较于化学杀虫剂具有其独特的优点:宿主专一不伤害人畜,不会产生残余物污染环境,不会引起昆虫产生抗性。但杆状病毒杀虫剂具有杀虫谱太窄、杀虫速度缓慢等缺点,因此其应用受到限制。深入了解病毒基因的结构和功能,进一步研究其致病机理,为遗传改良杆状病毒杀虫剂以扩大其杀虫谱和提高其杀虫效率提供了重要的理论指导。
SpltMNPV属杆状病毒科核多角体病毒属,基因组全长139 342 bp,含有141个开放阅读框(Open Reading Frame,ORF)[12]。到目前为止,在所有已测序的杆状病毒基因组中发现38个核心基因[13-16],SpltMNPV ORF75(Splt75)是杆状病毒核心基因之一,其在苜蓿丫纹夜蛾核多角体病毒(Autographa californica multiple nucleopolyhedrovirus,AcMNPV)中的同源基因ORF78(Ac78)在杆状病毒的生活周期中具有重要作用[17],但其作用机理未知。蛋白结构与功能息息相关,本研究选取到目前为止尚未见研究报道的Splt75作为研究对象,利用生物信息学方法系统分析Splt75基因及其编码蛋白Splt75的结构,为进一步揭示Splt75的功能和作用机理奠定基础。
1 材料和方法
1.1 材料
SpltMNPV G2毒株,由中山大学有害生物控制与资源利用国家重点实验室赠送提供,大肠杆菌(Escherichia coli)DH5α为本实验室保存。
pMD18-T Vector、Taq酶,购自TaKaRa公司;氨苄青霉素,购自Sigma公司;限制性内切酶,购自New England BioLabs公司;PCR产物纯化试剂盒、常规质粒提取试剂盒,购自Omega公司;其他试剂均为国产分析纯。
1.2 SpltMNPV基因组DNA的提取
参照文献[18]的方法,从SpltMNPV多角体中提取病毒基因组DNA。
1.3 PCR扩增及克隆
根据Genbank登录的SpltMNPV G2株基因组全序列[12](AF325155)中的Splt75基因全长序列,设计一对引物扩增Splt75基因,由广州擎科生物技术有限公司合成。上游引物PF:5'-GAGCTCATGAATTT GGACGTACCCTACGA-3',下游引物 PR:5'-GGATC CCTAAATGTACAAAGAGTCGGGAC-3',下划线部分分别为引入的SacI和BamHI酶切位点。以所提取的SpltMNPV基因组DNA为模板进行PCR扩增,回收PCR扩增产物,克隆入pMD18-T Vector,转化感受态DH5α,挑取并过夜培养阳性克隆,提取质粒,酶切鉴定,并在广州擎科生物技术有限公司进行测序,比较测序结果与Splt75基因序列。
1.4 生物信息学分析
使用DNASTAR软件推导出测序得到的Splt75基因序列编码蛋白Splt75的氨基酸序列,应用在线工具ExPASy ProParam(https://web.expasy.org/protparam)分析Splt75蛋白的分子式、氨基酸组成和相对分子量等理化性质。蛋白质的亲疏水特性预测使用ProtScale(https://web.expasy.org/protscale),蛋 白 质信号肽的检测使用SignalP(http://www.cbs.dtu.dk/services/SignalP),蛋白质结构域与模式的预测使用 TMHMM Server(http://www.cbs.dtu.dk/services/TMHMM)、MotifScan(https://myhits.isb-sib.ch/cgi-bin/motif_scan) 和 Psort(http://www.genscript.com/psort.html)等在线软件。利用NCBI Conserved Domain Search搜索工具搜索Splt75的保守区域。蛋白质二级结构利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl)在线软件进行分析,蛋白质三级结构的预测利用SWISS-MODEL在线工具(https://www.swissmodel.expasy.org)。
2 结果与分析
2.1 Splt75基因的克隆
以所提取的SpltMNPV基因组DNA为模板,从SpltMNPV基因组中PCR扩增出一条特异的363 bp左右的片段,克隆入pMD18-T载体,转化感受态DH5α,培养阳性克隆,提取质粒,用SacI和BamHI进行双酶切鉴定,获得与预期相符的363 bp的目的片段和2 692 bp的载体片段(图1),质粒命名为T-Splt75。T-Splt75的测序结果显示,pMD18-T载体中的插入片段与Genbank登录的SpltMNPV G2株基因组全序列(AF325155)中的Splt75基因全长序列完全一致,没有发生碱基突变。

图1 重组质粒T-Splt75的鉴定
2.2 Splt75蛋白的理化性质分析
通过DNASTAR软件和在线软件ExPASy ProParam (https://web.expasy.org/protparam) 分 析Splt75基因编码蛋白Splt75的理化性质。Splt75蛋白由120个氨基酸组成(图2),其中缬氨酸(Val)13个、亮氨酸(Leu)12个,含量相对较高,分别为10.8%和10.0%;共11个带负电荷氨基酸(Asp、Glu),占9.2%;12个带正电荷氨基酸(Arg、Lys),占10.0%;预测相对分子质量为13 137.16 Da,蛋白等电点为7.86,分子式为C583H942N160O174S5。Splt75蛋白的半衰期为30 h;不稳定指数为37.4(小于40),是稳定蛋白。
2.3 Splt75蛋白的疏水性分析
运用 ProtScale(https://web.expasy.org/protscale)在线软件分析Splt75蛋白的亲疏水性,结果如图3所示。Splt75蛋白第103位的精氨基(Arg)分值最低,为-3.189,第83位的亮氨酸(Leu)和84位的半胱氨酸(Cys)的分值最高,为3.989,在74~97氨基酸区域存在疏水区域,Splt75蛋白总体表现为疏水。
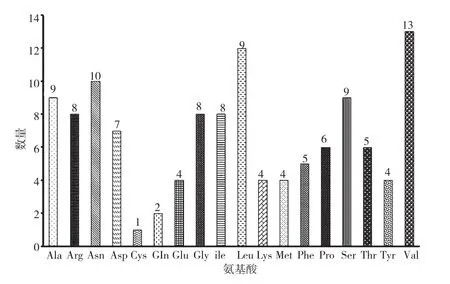
图2 Splt75的氨基酸组成

图3 Splt75的亲疏水性分析
2.4 Splt75的跨膜结构和信号肽预测
使用TMHMM(http://www.cbs.dtu.dk/services/TMHMM)预测Splt75蛋白的跨膜结构区,结果如图4所示。在Splt75蛋白的76~98氨基酸区域存在跨膜结构区。利 用 SignalP(http://www.cbs.dtu.dk/services/SignalP)在线预测显示Splt75蛋白无信号肽,Psort(http://www.genscript.com/psort.html)在线预测显示Splt75蛋白C末端无KDEL内质网滞留信号模序、内质网膜滞留信号模序和内质网回运信号类似模序(Endoplasmic Reticulum Retrieval Signals) (KKXX-like motif)。

图4 Splt75的跨膜结构区分析
2.5 Splt75的蛋白序列模式预测
运用Motif Scan(https://myhits.isb-sib.ch/cgi-bin/motif_scan)对Splt75蛋白进行分析,结果如表1所示。Splt75蛋白中可能含有1个N-糖基化位点(59~62),1个酪蛋白激酶Ⅱ磷酸化位点(30~33),1个蛋白激酶C磷酸化位点(32~34),1个N-豆蔻酰化位点(35~40),1个cAMP和cGMP依赖的蛋白激酶磷酸化位点(104~107),1个酰胺化位点(100~103)。
利用NCBI Conserved Domain Search搜索工具发现Splt75属于ORF78(Ac78)超家族成员(图5),该家族成员由AcMNPV ORF78及其同源的杆状病毒蛋白组成。

表1 Splt75蛋白序列模式分析结果

图5 Splt75的保守区域预测
2.6 Splt75的二级结构
运用在线软件SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl)分析Splt75蛋白的二级结构,结果如图6所示。Splt75二级结构的主要构成元件是α螺旋和无规则卷曲。其中α螺旋由48个氨基酸构成,占总氨基酸的40.00%,无规则卷曲由52个氨基酸构成,占总氨基酸的43.33%,18个氨基酸参与形成延伸链,占比15.00%,β转角由2个氨基酸构成,占比1.67%。

图6 Splt75二级结构预测
2.7 Splt75的三级结构
运用SWISS-MODEL在线工具(https://www.swissmodel.expasy.org)构建Splt75蛋白的三级结构模型,结果如图7所示。GMQE评分为0.16(质量评估的数字越大,结果越可靠),QMEAN评分为-2.25(基于不同几何特性的符合评估,分数≤-0.4表示一致性较差,模型可靠度较差)。

图7 Splt75三级结构预测
SpltMNPV的Splt75是杆状病毒核心基因,NCBI Conserved Domain Search搜索工具发现,Splt75属于ORF78(Ac78)超级家族成员,与Ac78的序列一致性为32%,表明Splt75可能具有与Ac78相似的功能。前期对Ac78的研究发现,Ac78蛋白是杆状病毒的一个多功能蛋白,主要参与BV的产生、多粒包埋ODV (multiple nucleocapsid-enveloped ODV,M-ODV)的形成和病毒在粉纹夜蛾幼虫体内起始初始感染的过程。Ac78的缺失导致BV和M-ODV的产量显著下降以及病毒在粉纹夜蛾幼虫体内起始初始感染能力的丧失,Ac78的氨基酸2~25和64~88区域在其所有旁系同源物(ortholog)中高度保守,是Ac78的关键区域[17]。
生物信息学是预测蛋白质功能以及分析蛋白质结构与功能之间关系的重要方法,在医学、生物技术等多个领域应用广泛[19]。通过生物信息学分析,Splt75蛋白由120个氨基酸组成,相对分子质量为13 137.16 Da,蛋白等电点为7.86,性质稳定,总体表现为疏水。Splt75蛋白无信号肽,存在跨膜结构区,无KDEL内质网滞留信号模序、内质网膜滞留信号模序和内质网回运信号类似模序(KKXX-like motif)。前期研究中通过生物信息学分析发现,在Ac78的C末端存在一个可能的内质网回运信号类似模序(KKXX-like motif),通过构建缺失Ac78 KKXX-like模序的重组病毒vAc78:del105-108研究发现,该模序在Ac78功能和在感染细胞中定位非必需[20],这与该模序在Ac78的同源蛋白中并不保守相一致。
蛋白质的翻译后修饰对调节蛋白质的活力与功能具有非常重要的作用[21]。通过对Splt75的蛋白序列模式预测发现,Splt75有N-糖基化、酪蛋白激酶Ⅱ磷酸化、蛋白激酶C磷酸、N-豆蔻酰化、cAMP和cGMP依赖的蛋白激酶磷酸化和酰胺化等多个翻译后修饰位点。在Ac78中,经预测可能存在2个N-糖基化和1个酪蛋白激酶Ⅱ磷酸化翻译后修饰位点(文中未显示),表明N-糖基化位点和酪蛋白激酶Ⅱ磷酸化位点在Splt75及其同源蛋白中相对保守,可能在Splt75及其同源蛋白的功能中具有重要作用。对Splt75的二级结构预测发现,α螺旋和无规则卷曲为Splt75的主要结构。α螺旋的存在有利Splt75蛋白结构的稳定,α螺旋占比40.00%,说明Splt75可能相对保守,这与Splt75是杆状病毒核心基因相符。无规则卷曲结构的存在区域可能是Splt75发挥功能的重要区域,无规则卷曲占比43.33%,说明Splt75可能在杆状病毒生活周期中具有重要作用。
3 结论
Splt75基因编码120个氨基酸,编码蛋白Splt75分子量为13 137.16 Da,蛋白理论等电点为7.86,分子式为C583H942N160O174S5,是稳定的疏水蛋白;Splt75蛋白中无信号肽,存在跨膜结构区,C末端无KDEL内质网滞留信号模序、内质网膜滞留信号模序和内质网回运信号类似模序;可能含有1个N-糖基化位点、1个酪蛋白激酶Ⅱ磷酸化位点、1个蛋白激酶C磷酸化位点、1个N-豆蔻酰化位点、1个cAMP和cGMP依赖的蛋白激酶磷酸化位点以及1个酰胺化位点;属于ORF78(Ac78)超家族成员;Splt75的二级结构以α螺旋和无规则卷曲为主要构成元件。本研究结果可靠,为进一步研究Splt75在杆状病毒生活周期中的功能和作用机理提供了理论基础。
猜你喜欢
军事文摘(2022年16期)2022-08-24
波谱学杂志(2022年1期)2022-03-15
昆明医科大学学报(2022年1期)2022-02-28
临床肝胆病杂志(2021年7期)2021-12-26
今日农业(2021年11期)2021-08-13
现代临床医学(2021年1期)2021-01-26
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
分析化学(2017年12期)2017-12-25
安徽医科大学学报(2015年9期)2015-12-16
