抑郁症发病与神经营养因子异常研究进展
2020-10-28 09:54王惠芹王真真林美妤陈乃宏
中国药理学通报 2020年10期
王惠芹,王真真,林美妤,陈乃宏
(1. 湖南中医药大学药学院,湖南省中药饮片标准化及功能工程技术研究中心,湖南 长沙 410208;2.中国医学科学院药物研究所,神经科学中心,北京 100050)
抑郁症(depression)是一种以心境低落、思维迟缓、动力减退甚至悲观厌世、企图自杀为主要临床症状的精神性疾病,抑郁症的发病过程涉及生理、心理和社会环境等多因素同时作用,以致其发病机制迄今尚未完全阐明。神经营养因子是神经可塑性的有效调节因子,其活性降低会导致细胞脆弱性增加甚至死亡[1]。抗抑郁药物治疗后神经营养因子分泌增加,从而促进神经元的存活,保护神经元免受应激损伤。神经营养因子与抑郁症关系的研究表明神经营养因子有望成为抗抑郁治疗的一类有效靶点。通过重点总结脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)、血管内皮生长因子(vascular endothelial growth factor,VEGF)、胶质细胞源性神经营养因子(glial cell-derived neurotrophic factor,GDNF)、神经生长因子(nerve growth factor,NGF)和其他神经营养因子及其相关基因与抑郁症发病的关系,有助于我们更好地了解神经营养因子对抑郁症发病的重要影响,促进我们将神经营养因子作为有效的药物干预抑郁症的候选靶点,为抑郁症患者制定有效的个体化治疗方案。
1 BDNF
BDNF是神经营养因子家族的重要成员,1982年由Barde等首次在猪脑中发现[2]。BDNF主要在神经元中合成并广泛存在于中枢神经系统,参与神经可塑性和对应激性神经损伤的修复,影响5-羟色胺(5-hydroxytryptamine,5-HT)信号转导,增加脑组织中5-HT水平,防止5-HT神经元退化。越来越多的研究表明,BDNF与抑郁症的发生、发展和治疗有关,是目前抑郁症相关神经生物学领域研究最多的神经营养因子。研究学者不断探索BDNF活性与含量的变化和抑郁症的发生与结果之间的关系,Guilloux等[3]检测到女性重度抑郁症(major depressive disorder,MDD)患者的杏仁核中BDNF功能降低;Lee等[4]表明有抑郁情绪、抑郁症复发和自杀企图的患者体内血浆BDNF水平降低。此外,Fukumoto等[5]表明(2R, 6R)-羟基去甲氯胺酮[(2R, 6R)-HNK]——有效抗抑郁候选药物之一氯胺酮的代谢产物,在抑郁症动物模型中产生快速和持续的抗抑郁作用而没有氯胺酮的副作用,而且其抗抑郁作用需要通过BDNF的释放介导,从而增强内侧前额叶皮质的突触传递功能,这一重大发现再次证明BDNF在抗抑郁治疗中不可或缺的作用。
原肌球蛋白相关激酶B (tropomyosin-related kinase B,TrkB) 是BDNF的特异性受体,最近研究表明,烟碱的抗抑郁作用需要通过BDNF-TrkB信号的激活[6],氯胺酮通过上调BDNF-TrkB信号改善术后抑郁症状[7]。然而BDNF与抑郁症关系的结果仍不一致,Wook Koo等[8]指出在中脑腹侧背盖区(ventral tegmental area,VTA)-伏隔核( nucleus accumbens,NAc)通路中,NAc中BDNF蛋白表达水平升高诱发抑郁样行为,在VTA中局部敲除BDNF基因降低抑郁样表型,证明了从VTA转运至NAc的BDNF信号诱导抑郁易感性。药物阻断NAc中BDNF-TrkB信号,减少VTA中BDNF水平,可以防止社交应激诱发的抑郁样行为,可见不同脑区的BDNF作用机制不同。
位于染色体2和染色体11上的BDNF基因与MDD和抑郁易感性相关。最受关注的BDNF的遗传标记是在196(G/A)核苷酸上的单核苷酸多态性,其中66号密码子中一个缬氨酸被蛋氨酸取代,通常被称为val66met(rs6265),影响BDNF的加工和分泌。Fukumoto等[5]表明,将BDNF-val66met等位基因敲入内侧前额叶皮质或向其内注射微量抗BDNF中和抗体会阻断抗抑郁药物对小鼠的治疗作用;当增加内侧前额叶皮质BDNF释放会导致下游TrkB的激活,从而增加内侧前额叶皮质锥体神经元的突触功能,逆转应激和抑郁的损伤,证实BDNF-val66met(rs6265)基因多态性与抑郁样行为和抗抑郁药物抵抗密切相关。尽管有强有力的文献支持BDNF基因的val66met多态性与抑郁症之间的关联,但与大多数其他抑郁症候选基因类似,其实验结果并不一致。Schumacher等[9]对465名MDD患者进行基因检测,未发现MDD与val66met有明显相关性。BDNF基因与其他基因对抑郁症发病机制有交互作用,且受环境的影响,应进一步探究其潜在的抑郁症发病机制,深入研究BDNF基因与抑郁症的关系。
BDNF与抑郁症关系的研究结果尚不一致,仍有如下问题亟待解决:(1)不同脑区BDNF的变化水平和作用机制不同,杏仁核[3]和内侧前额叶皮质[5]中BDNF水平抑制抑郁,而VTA-NAc通路中BDNF水平促进抑郁[8],各个脑区的BDNF水平变化对抑郁症发病有何种影响,尚需进一步研究;(2)不同的酶联免疫吸附试验方法或取样管可能导致BDNF测定值有差异,而目前测量BDNF的水平没有唯一而确定的方法,未来需要探索更稳定且精准的BDNF测量方法;(3)不同脑区BDNF基因转录水平有很大差异,且它在大脑中的含量非常低,阻碍了针对抑郁症相关脑区中BDNF表达的神经元的新药物发展。最新研究揭示可用绿色荧光蛋白(green fluorescent protein,GFP)作为识别成年小鼠大脑中BDNF mRNA水平的替代标记物,可从BDNF-GFP构建物取代BDNF基因的小鼠系中提取单个细胞进行RNA测序[10]。
2 VEGF
VEGF是一种重要的神经营养因子,于1984年首次由Chen等[11]在胎牛视网膜中提取分离得到。VEGF参与血管生成的生理病理过程[12],在中枢和周围神经系统具有神经营养和神经保护潜能。此外,VEGF是内皮细胞和神经元的一种有效的有丝分裂原和生存因子,是突触传递的调节剂[13]。研究表明VEGF在海马神经发生和抑郁症病理生理学中起重要作用,抑制神经细胞中VEGF受体2的表达会损害海马依赖性突触可塑性和情感记忆的巩固[14]。目前临床上对VEGF与抑郁症关系的研究结果尚不一致,MDD患者血浆中的VEGF水平与正常对照组相比均降低[15],然而,有研究表明MDD患者体内血浆VEGF水平有升高趋势[16],可能是由于诸如性别、年龄、体重指数等环境因素未充分评估而导致结果相矛盾。
人类VEGF基因位于6号染色体(6p21.3)上,有多种基因多态性,这种多态性可能与抑郁易感性以及抑郁症发病有关。Xie等[17]通过一项437名抑郁症患者和477名健康志愿者的病例对照研究,探讨4种与VEGF相关的单核苷酸多态性(rs10738760、rs6921438、rs6993770和rs4416670)与抑郁症之间的潜在联系,结果发现单核苷酸多态性rs4416670与抑郁风险增加相关(OR=1.60,P=0.010),证明了VEGF基因与抑郁症有关。另有研究表明MDD组与对照组VEGF基因rs2010963位点各基因型频率之间的差异有统计学意义,说明VEGF基因rs2010963位点基因多态性与抑郁症的产生或结果有关[18]。此外,该实验还发现rs2010963位点上携带C等位基因(CC+CG)的个体在暴露于高强度负性生活压力事件时罹患抑郁症的可能性更大,VEGF rs2010963多态性与环境的相互作用增加了MDD发病风险[18],因此环境因素在抑郁症的发病过程中也起着重要的作用。这一新的关联揭示了抑郁症新的分子机制,表明VEGF基因在抑郁症发展中的潜在作用。
通过VEGF与抑郁症关系的研究,可以发现如下几个问题:(1)尽管BDNF是目前MDD神经生物学领域研究最多的神经营养因子,但VEGF对抑郁症发病的影响不容小觑,应深入探讨VEGF与抑郁症之间的关系;(2)目前VEGF与抑郁症相关性的研究结果尚不一致,未来的研究应结合性别、年龄、体重指数等环境因素全面研究VEGF对抑郁症发病机制的影响;(3)VEGF及其基因异常为阐明抑郁症的发病机制提供了新的视角,进一步的研究将有可能使VEGF成为治疗抑郁症的新的分子靶点。
3 GDNF
GDNF是转化生长因子β家族的一种神经营养因子,由Lin等[19]于1993年首次分离纯化并命名。GDNF在整个大脑中广泛表达,调节去甲肾上腺素能和γ-氨基丁酸能系统,是影响中脑多巴胺能神经元发育、存活和活性的最有效的神经营养因子之一,并保护神经元和胶质细胞免受氧化应激的损害,在海马细胞的发育和功能完整性中起着重要作用。Hamidi 等[20]对MDD患者的尸检研究发现MDD患者杏仁核GDNF减少,表明较低的杏仁核GDNF可能参与MDD的病理生理学。Zhang等[21]发现MDD患者治疗前血清GDNF明显低于健康对照组,急性发作期外周GDNF合成及释放减少,抗抑郁药物可通过增加GDNF水平达到治疗效果,并以此推测GDNF水平的下降可能是抑郁症状态的一种生物标志。有研究表明中枢GDNF信号也可能是抗抑郁治疗的新靶点[22]。
Uchida等[23]使用应激易感BALB和应激抵抗B6两种基因不同的小鼠品系进行研究,通过其对慢性不可预知温和应激(chronic unpredictable mild stress, CUMS)表现出的不同行为反应探究腹侧纹状体GDNF基因的表观遗传学差异状态如何调节对CUMS的敏感性和适应性,结果发现CUMS导致BALB小鼠腹侧纹状体内甲基CpG结合蛋白2-组蛋白去乙酰化酶2复合物增多,并与GDNF启动子结合抑制GDNF转录。此外,GDNF启动子CpG位点的DNA甲基化增多,降低GDNF表达,从而诱发BALB小鼠的抑郁样行为,表明GDNF启动子的组蛋白修饰和DNA甲基化促使小鼠产生抑郁样行为,为腹侧纹状体中GDNF基因的表观遗传差异状态如何影响应激的行为反应差异机制研究提供了一定的见解。之后多项研究再次表明GDNF启动子DNA甲基化水平的上调[24]和组蛋白去乙酰化酶2水平升高[25]参与大鼠抑郁样行为的发生。GDNF基因位于5p13.1-p13.3,有实验研究了218名中国汉族抑郁症患者中与GDNF相关的6个单核苷酸多态性(rs2910709、rs2973050,rs884344,rs2910702,rs2216710,rs3812047)与抑郁症的关系,结果发现rs884344 A/C和C/C基因型(OR=1.601)以及C等位基因(OE=1.532)与其他基因型以及等位基因相比,与抑郁症的风险增加显著相关[26],表明GDNF基因是抑郁症的新易感性基因。
通过以上GDNF与抑郁症发病关系的研究,可以发现:(1)外周GDNF与中枢GDNF水平对抑郁症发病的影响机制是否一致有待验证;(2)GDNF对血脑屏障的穿透性很差,未来的研究可考虑测量不同程度的抑郁症患者外周和中枢GDNF水平以进一步检验外周和中枢GDNF是否相关。
4 NGF
NGF是神经营养因子家族的重要成员,于1956年由Levi-Montalcini首次分离得到[27],主要产生于皮层、海马和下丘脑,也存在于外周神经系统和免疫系统。NGF在神经元存活、神经可塑性和学习记忆中起着重要作用,对应激反应至关重要,参与神经-内分泌-免疫(neuro-endocrine-immune,NEI)系统的调节,而神经可塑性和NEI系统稳态的改变在抑郁症的病理生理学中起重要的作用,因此NGF涉及抑郁症的发病机制。Wiener等[28]评估有或无自杀风险(suicide risk, SR)的MDD患者NGF血清水平的变化并探讨NGF在抑郁症和自杀风险的病理生理学中的可能作用,结果发现与对照组相比,MDD组和MDD+ SR组的NGF血清水平显著降低,然而MDD组和MDD+ SR两组之间的NGF血清水平没有差异,由此指出NGF可能是MDD的一种生物标志物,而其抑郁程度无法通过血清NGF水平降低程度判别。人类早期的不良经历与罹患诸如焦虑和抑郁症等精神疾病的风险增加有关,如母爱剥夺增加抑郁症的易感性,是一种潜在的抑郁症动物模型。Cirulli等[29]发现雌性恒河猴幼崽在经历母爱剥夺后外周NGF水平降低,表现出抑郁样行为,而雄性恒河猴幼崽在经历母爱剥夺后外周NGF水平升高,对应激产生积极应对反应,表明NGF在脑功能和神经保护中发挥重要作用且受应激影响,是影响脑功能变化中不良事件转导的良好候选因子,是雌、雄性恒河猴幼崽对母性剥夺后不同反应的神经内分泌标志物。
NGF基因位于染色体1p13.1上,Yeh等[30]为研究NGF基因多态性是否与抗抑郁药物治疗MDD患者的疗效相关,对经过八周抗抑郁治疗的抑郁症患者体内NGF基因的7个单核苷酸多态性(rs6330, rs2254527, rs6678788, rs12760036, rs7523654, rs17033692, rs11102929)进行了基因分型,结果在3个NGF单核苷酸多态性(rs12760036, rs7523654和rs17033692)中观察到缓解型相较非缓解型基因型频率显著增加。单倍型分析显示C/C/C单倍型(rs2254527-rs6678788-rs12760036)与较高的缓解率相关,而C/C/A单倍型与较低的缓解率相关,证明了NGF基因多态性与抗抑郁治疗后的缓解率相关。
根据以上NGF与抑郁症关系的研究,可以认识到:(1)Wiener等[28]评估伴有或不伴有SR的MDD患者NGF血清水平的变化,发现自杀风险的存在并不影响抑郁症患者NGF的血清水平,这表明NGF可能与MDD的诊断有关,而与症状的严重程度无关;(2)研究NGF基因多态性与抑郁症发病和抗抑郁药物疗效关系时,所选的单核苷酸多态性未覆盖整个NGF基因,其他单核苷酸多态性与抑郁症发病是否有关仍有待研究。
5 总结与展望
综上所述,神经营养因子长期以来被认为是结构可塑性的关键介质,大量研究支持其在抑郁症发病和抗抑郁治疗中的作用,并将其作为有效的药物干预抑郁症的候选靶点。神经营养因子如BDNF、VEGF、GDNF和NGF等与抑郁症发病相关,通过总结抑郁症发病与神经营养因子及其基因异常的关系(Table 1),有利于为抑郁症的早期诊断和治疗提供新型靶点。然而神经营养因子与抑郁症关系的研究尚未取得一致性结果,原因可能包括实验设计、实验环境测量指标、抑郁样行为和程度测量方法以及研究对象的种族、生长环境背景等多方面的差异。目前BDNF与抑郁症关系的研究较为成熟,而近年来VEGF、GDNF、NGF等神经营养因子与抑郁症关系的研究也越来越多,并结合了自杀风险、年龄以及是否经过抗抑郁药物治疗等多方面因素进行探究,支持了“神经营养因子假说”和“神经可塑性假说”在抑郁症发病机制中的应用。此外,胰岛素样生长因子-1有助于神经元的成熟并调节成年神经发生,粒细胞集落刺激因子可提高细胞存活率和活力、增加海马神经元可塑性,在抑郁症的病理生理和抗抑郁治疗中起干预作用。成纤维细胞生长因子、转化生长因子、表皮生长因子和血小板源生长因子等神经营养因子均有可能与抑郁症发病存在一定的关系,但目前的研究相对较少,未来还有更多的神经营养因子与抑郁症的关系有待我们进一步发现与研究。目前,识别能够帮助诊断或监测抑郁症病情进展和治疗效果的生物标志物仍然是抑郁症研究领域的一个重点目标。未来研究可通过采用多基因交互-环境设计,考察研究对象种族、性别、年龄、家庭环境和社会环境的调节作用,全面探索潜在的抑郁症发病机制,进一步推进和完善有关神经营养因子及其基因异常与抑郁症发病机制的研究。
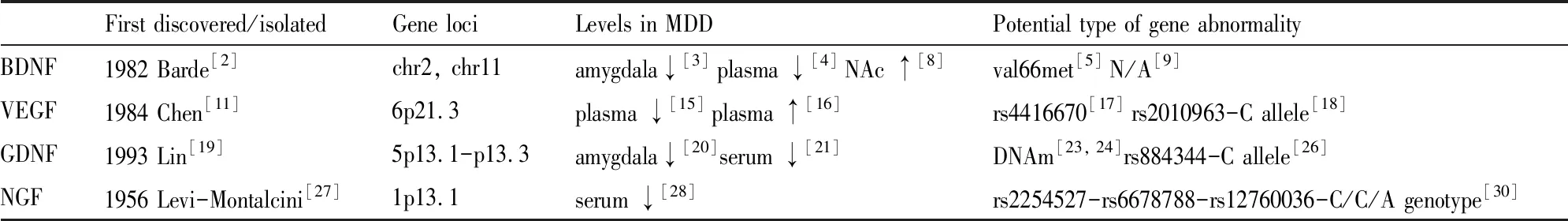
Tab 1 Discovery and relationship between neurotrophins and pathogenesis of depression
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
世界科学技术-中医药现代化(2022年3期)2022-08-22
世界科学技术-中医药现代化(2022年3期)2022-08-22
保健医苑(2022年4期)2022-05-05
昆明医科大学学报(2022年2期)2022-03-29
中老年保健(2021年5期)2021-12-02
昆明医科大学学报(2021年6期)2021-07-31
昆明医科大学学报(2021年3期)2021-07-22
电子产品世界(2021年8期)2021-01-16
中国疼痛医学杂志(2017年8期)2017-01-11