
p62 基因缺失对人脂肪间充质干细胞成脂的促进作用
2020-10-21 23:49:36曾瑞霞张轶博
吉林大学学报(医学版) 2020年5期
曾瑞霞,张轶博
(1.锦州医科大学基础医学院解剖学教研室,辽宁 锦州 121001;2.锦州医科大学基础医学院病原生物学教研室,辽宁 锦州 121001)
肥胖发生的根本原因是脂肪细胞体积变大和数量增多,而后者很大程度上是由于脂肪分化异常所致[1-3]。脂肪间充质干细胞(adipose-derived stromal cells,ADSCs)作为脂肪细胞形成的源头,其增殖能力和分化潜能直接决定了成熟脂肪细胞的数量,影响脂肪组织的结构与构成。p62 是自噬-溶酶体系统中重要的中介分子[4-5],p62 基因敲除的小鼠发生肥胖和胰岛素抵抗,白色脂肪组织中脂肪积累增加,脂肪内脂质的合成增加[6]。自噬相关基因7(autophagy-related gene 7,Atg7)敲除的小鼠以及Atg7 缺失的小鼠3T3-L1 前脂肪细胞的研究显示:脂肪组织中总脂肪堆积量和脂滴数量减少,且脂滴体积缩小,小鼠3T3-L1 前脂肪细胞分化的成熟脂肪细胞内脂滴数量减少,表明自噬被抑制后可降低甘油三酯积累,抑制脂肪细胞分化。线粒体自噬是一种选择性自噬,即自噬泡膜上有识别受损线粒体的特异受体,并对其针对性地进行降解,这对于维持线粒体正常的数量和功能非常重要。目前,关于脂肪分化的研究多以小鼠3T3-L1前脂肪细胞为模型,本研究选用人脂肪间充质干细胞(human adipose-derived stromal cells,hADSCs)作为研究对象进行脂肪分化研究,该细胞与小鼠3T3-L1 细胞比较更接近人的生物学状态,对疾病的预防和治疗更具有指导意义。肥胖和高脂饮食有密切关联[7-8],而油酸(oleic acid,OA)是动物油和植物油中常见的游离脂肪酸,也是血液游离脂肪酸(free fatty acid,FFA)含量最多的成分,将其替代3-异丁基-1-甲基黄嘌呤(3-isobutyl-1-methylxanthine,IBMX)进行成脂诱导,可在一定程度上模拟人体的生理作用模式。p62 是否通过调节脂肪细胞的分化而影响肥胖的发生尚未见文献报道,故本研究构建p62 shRNA 慢病毒载体,探讨p62 基因缺失对hADSCs 成脂能力的影响以及与自噬之间的关系。脂肪分化过程中伴随着脂滴的形成及甘油三酯的合成,而线粒体是脂肪酸氧化的核心场所,因此在明确自噬在hADSCs 成脂过程中的基本作用后,进一步探讨线粒体自噬与p62 基因在hADSCs 成脂中的作用及关系,为更深入阐明人脂肪分化的分子机制和寻找新药的作用靶点提供新思路。
1 资料与方法
1.1 研究对象、主要试剂和仪器脂肪组织取自3 例大腿皮下脂肪抽吸术者(锦州医科大学附属第三医院整形外科),供者均为25~35 岁的健康女性,并对实验知情同意。单丹磺酰戊二胺(monodansylcadaverine,MDC)(大连宝生物工程有限公司),细胞计数试剂盒(cell counting kit-8,CCK-8)(美国Invitrogen 公司),Mitotraker Red 探针(美国Invitrogen 公司),一抗兔抗人β-actin、微管相关蛋白1 轻链3(microtubule-associated protein 1 light chain 3,LC3)、CCAAT 增强子结合蛋白 α(CCAAT/enhancer binding protein alpha,CEBPα)、p62 单克隆抗体和羊抗兔IgGHRP 二抗(美国Sigma 公司)。DP71 型正置荧光显微镜(日本Olympus 公司),PRISM 7300 型实时荧光定量PCR 仪(美国ABI 公司)。
1.2 p62 基因的shRNA 引物合成和慢病毒载体构建根据GenBank 公布的人p62 基因核苷酸序列(NM_001142299),人p62 基因特异性shRNA 靶序列(5'-GCAGATGAGGAAGATCGCCTT-3')的设计、合成以及慢病毒载体的构建由上海吉凯基因化学技术有限公司完成。
1.3 hADSCs 的分离和培养将皮下脂肪抽吸液装入50 mL 离心管,0.1%的Ⅰ型胶原酶进行消化,于37℃水浴箱作用1 h;加入等体积的完全培养基终止消化,1 500 r·min-1离心10 min,弃上清,留沉淀;将沉淀使用7 mL 低糖DMEM 进行吹打混匀,200 目网过滤,收集滤液;1 000 r·min-1离心5 min,弃上清,留沉淀;L-DMEM 吹打混匀沉淀,将其移入75cm2的培养瓶中,放入5% CO2、37℃恒温细胞培养箱中进行常规培养。
1.4 OA诱导hADSCs的脂肪分化采用80 μmol·L-1OA、10 mg·L-1胰岛素和1 μmol·L-1地塞米松在hADSCs 成脂的全程进行持续诱导10 d,镜下观察成脂情况。
1.5 CCK-8 实验绘制hADSCs 生长曲线将对照组和p62 基因缺失组hADSCs 接种于96孔细胞培养板中,每孔1×103个细胞,并于接种后0~7d检测细胞活性,每孔加入10μLCCK-8溶液,于37℃、5% CO2孵箱中继续培养2 h,使用分光光度计在450 nm 波长处测定吸光度(A)值,以A值为纵坐标,培养天数(d)为横坐标绘制细胞生长曲线。
1.6 Western blotting法检测hADSCs中p62、C/EBPα 和LC3Ⅱ/LC3Ⅰ蛋白表达水平取第3 代对数生长期hADSCs 接种于6孔水平板中,待细胞达50%融合后加入重组慢病毒感染。24h后PBS缓冲液冲洗3次,待细胞继续生长2d,诱导10d,进行尼罗红染色。于接种后10d采用Western blotting法检测hADSCs中p62、C/EBPα和自噬相关蛋白LC3Ⅱ/LC3Ⅰ的表达水平。
1.7 MDC 荧光染色检测自噬囊泡将P3 代hADSCs 接种到放有盖玻片的24 孔细胞培养板上进行成脂诱导,3d后进行染色观察。将细胞用PBS 缓冲液清洗3次后,换为含MDC的培养基37℃孵育10min,PBS缓冲液洗涤2次,取出盖玻片,用紫外光激发,于倒置荧光显微镜下观察和摄片。
1.8 Mitotracker Red 荧光共定位检测hADSCs 中线粒体自噬活性将P3 代hADSCs 接种到6 孔细胞培养板上进行成脂诱导,3d后加入200 nmol·L-1的MitotrackerRed进行染色15 min,PBS 缓冲液冲洗3 次,4%多聚甲醛固定15 min。加入2 mL 0.1%Trition X-100 作用20 min;10%山羊血清封闭30 min;加入兔抗人LC3多克隆抗体,4℃过夜;加入Alexa Fluor 鼠抗兔IgG 二抗,避光,37℃孵育1 h。将细胞置于荧光显微镜下观察并采集图像,计数各组含脂滴细胞数。
1.9 统计学分析采用SPSS 17.0统计软件进行统计学分析。各组hADSCs细胞增殖活性,MDC染色阳性细胞数,各组hADSCs中p62、C/EBPα和LC3Ⅱ/LC3Ⅰ蛋白表达水平均符合正态分布,以表示,多组间样本均数比较采用单因素方差分析,组间两两比较采用SNK-q检验。以P<0.05 为差异有统计学意义。
2 结果
2.1 hADSCs 的形态表现分离培养的hADSCs接种24h后可见细胞贴壁,呈短梭形。接种4d后,细胞开始增殖,并充分伸展,呈多角形或梭形。传代后细胞生长速度加快,呈漩涡状排列(图1)。
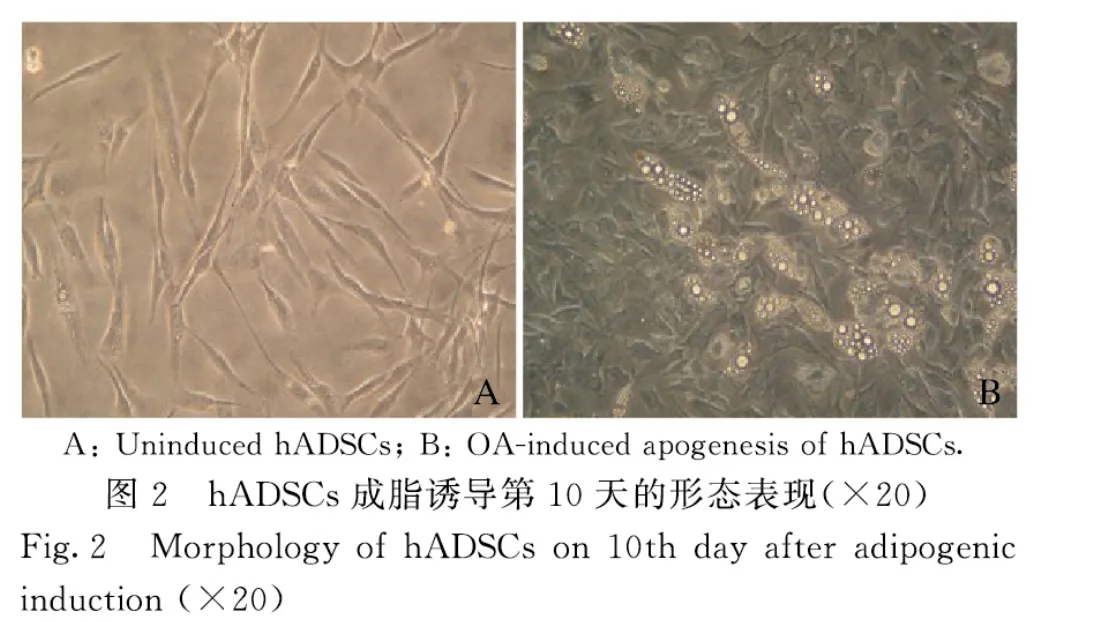
2.2 光镜下观察OA 诱导hADSCs 成脂后脂滴的形成选取80 μmol·L-1OA 联合地塞米松和胰岛素对hADSCs 进行成脂诱导10 d,光镜下观察到细胞内有大量脂滴形成(图2)。
2.3 hADSCs 生长曲线将慢病毒shp62 转染细胞24 h 后,更换培养液继续培养,结果显示:细胞形态未发生明显变化。采用CCK-8 比色法绘制生长曲线(图3),结果显示:在培养7 d 内,生长曲线呈“S”形,但p62 基因缺失组生长曲线较低,细胞增殖缓慢;在培养5 和6 d 后p62 基因缺失组细胞增殖活性较对照组明显降低(P<0.05 或P<0.01)。
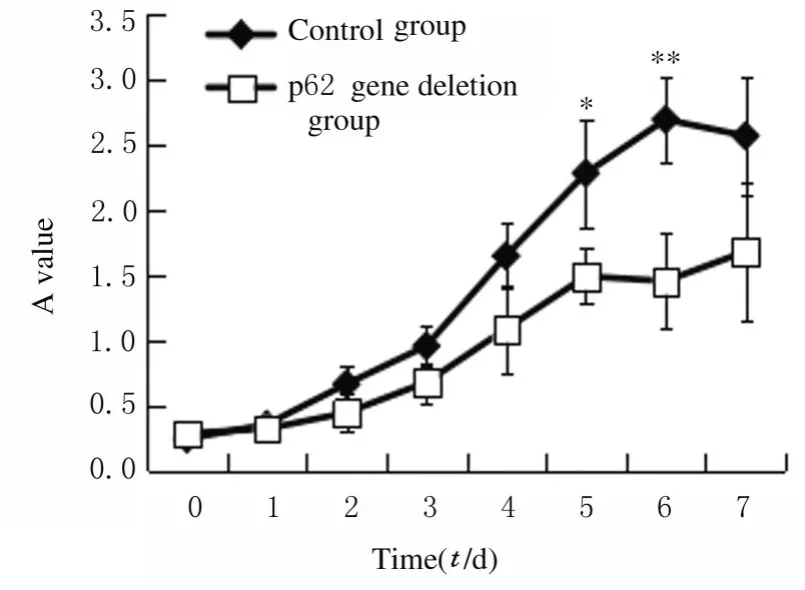
图3 2 组hADSCs 的生长曲线Fig.3 Growth curves of hADSCs in two groups
2.4 尼罗红染色结果和hADSCs 中C/EBPα 蛋白表达水平与对照组比较,p62 基因缺失组hADSCs 诱导脂肪分化至第3 天时,细胞形态变圆,细胞内已出现微小的脂滴,而对照组诱导第5 天才有脂滴出现;诱导分化至8~10 d,脂滴更丰富,细胞进一步变大变圆,尼罗红染色显示有丰富的脂滴,尼罗红脂滴染色阳性细胞数增多(图4)。Western blotting 法检测结果显示:p62基因缺失组C/EBPα 蛋白表达水平(1.40±0.10)较对照组(0.80±0.10)明显升高(P<0.01)(图5)。
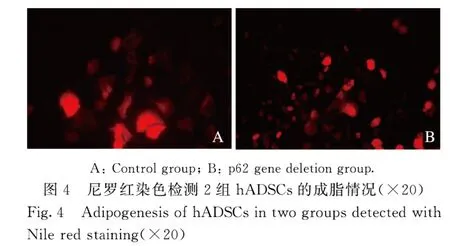
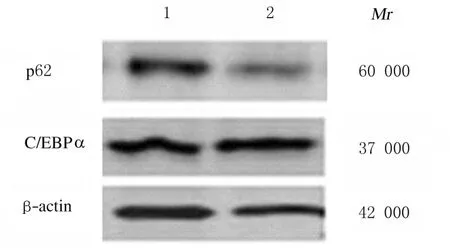
图5 Western blotting 法检测2组hADSCs中C/EBPα 蛋白表达电泳图Fig.5 Electrophoregram of expressions of C/EBPα protein in hADSCs in two groups detected by Western blotting method
2.5 MDC 染色结果和hADSCs 中LC3Ⅱ/LC3Ⅰ蛋白表达水平MDC 染色结果显示:对照组MDC荧光呈弥散分布,p62 基因缺失组为点状聚集,胞浆内荧光颗粒显著增多,荧光强度增加(图6),MDC 染色阳性细胞数(26.13±1.24)较对照组(16.34±1.42)明显增加(P<0.05)。Western blotting 法检测结果显示: p62 基因缺失组hADSCs 中LC3Ⅱ/LC3Ⅰ蛋白表达水平(0.80±0.10)较对照组(0.33±0.15)明显升高(P<0.01)(图7)。


图7 Western blotting 法检测2 组 hADSCs 中 LC3 Ⅱ/LC3Ⅰ蛋白表达电泳图Fig.7 Electrophoregram of expressions of LC3 Ⅱ/LC3 Ⅰproteins in hADSCs in two groups detected by Western blotting method
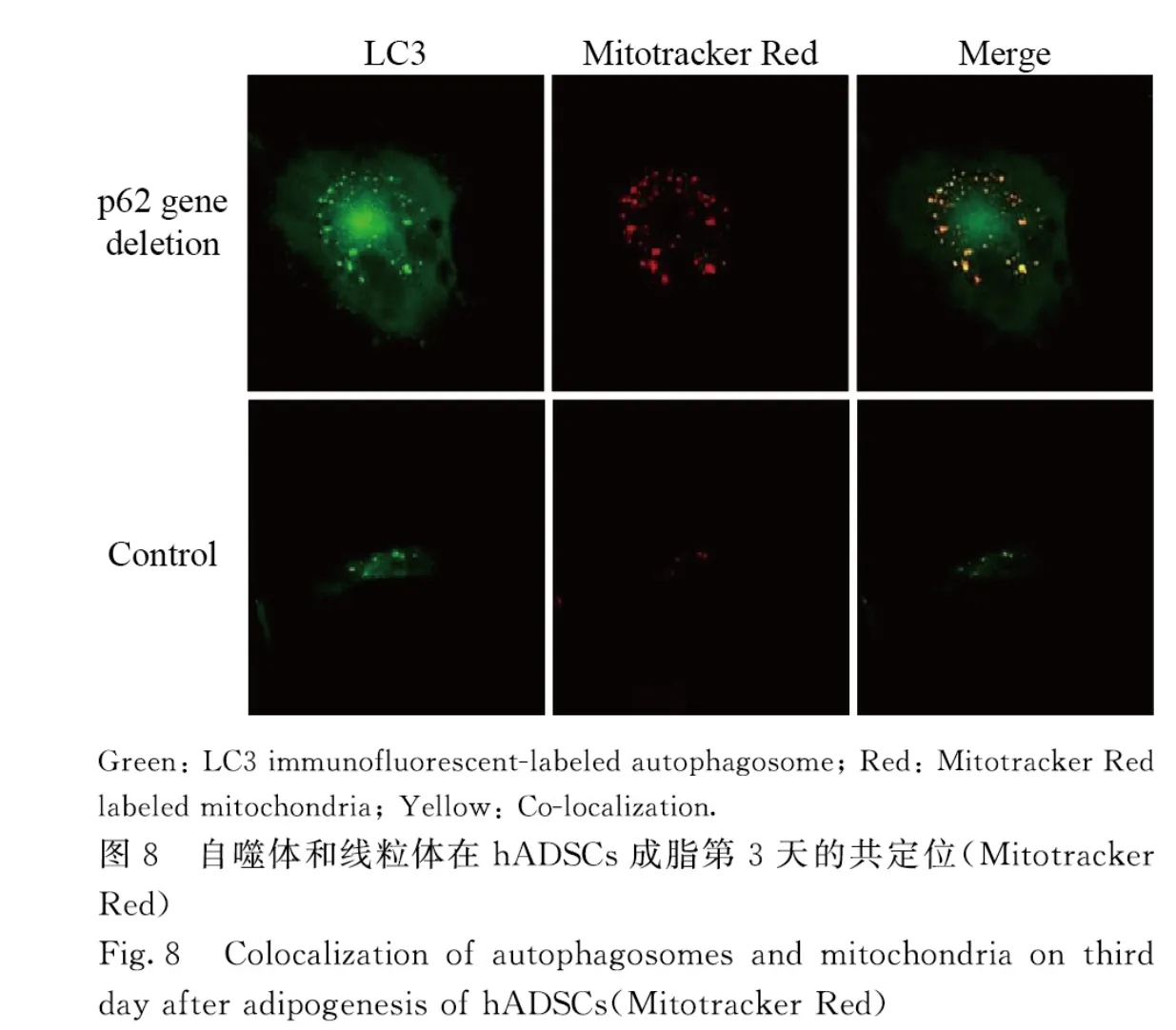
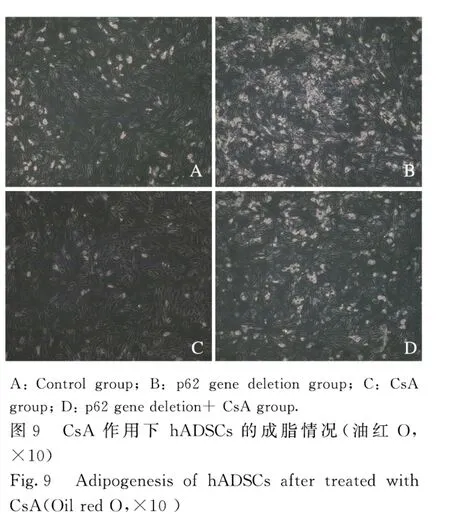
2.6 Mitotracker Red 荧光共定位检测hADSCs 中线粒体自噬活性与对照组比较,在分化3 d,p62基因缺失hADSCs 中LC3 的点状化和颗粒化增多,与线粒体的共定位现象增加(图8)。在分化10 d,光镜下计数含脂滴细胞数(图9),对照组为(26.67±1.53)个,p62 基因缺失组为(87.33±2.52)个,环孢菌素 A(cyclsporin A,CsA)组为(14.67±2.52)个,p62 基因缺失+CsA 组为(32.33±2.51)个。当p62 基因缺失引起线粒体自噬的增加被CsA 抑制后,其促进成脂的作用也被抑制,降低到接近对照组的水平。
3 讨论
p62 基因敲除小鼠出现肥胖和胰岛素抵抗,基础脂水解变慢,并在其脂肪组织中发现脂滴数量增多,甘油三酯合成增加,脂肪细胞体积增大。本研究将慢病毒shp62 转染至hADSCs 中,内源p62 基因沉默后,再进行成脂分化,结果显示:尼罗红染色阳性细胞数明显增多,调节成脂的关键分子C/EBPα 的蛋白表达水平明显升高,表明p62 基因缺失明显增强了hADSCs 的成脂能力。干细胞的自我复制能力,也是干细胞维持“稳态”的一种体现,而p62 基因缺失降低hADSCs 的自我复制能力,在一定程度上表明干细胞维持稳态的能力下降,可能趋向分化。
鉴于p62 作为自噬发生的重要中介物质,以及自噬在肥胖中的研究,本研究探讨p62 基因与自噬在hADSCs 成脂中的相互作用,MDC 染色及LC3Ⅱ/LC3Ⅰ蛋白检测结果显示:p62 基因的缺失在促进成脂过程中激活了自噬。研究[4]显示:如果p62 基因缺失,与p62 基因参与介导有关的自噬将降低;而本研究结果则显示:hADSCs 中p62 基因缺失导致自噬活性增强,与报道[4]结果相反,推测其可能原因:由于p62 基因也可介导泛素蛋白酶体系统(UPS)对小分子蛋白的降解,因此p62 基因缺失后,很多由p62 基因介导的小分子物质不能被UPS 清除,在细胞内大量聚集,后者可通过其他中介分子形成复合体,进而通过自噬进行降解,因此细胞的自噬活性增强。
研究[9-10]显示:线粒体在维持干细胞自我更新和定向分化中具有重要作用,当干细胞走向分化时,线粒体逐渐“成熟”,数量增加,功能增强。成脂过程中线粒体增加可能与以下2 个因素有关:①细胞分化过程需要线粒体提供足够的ATP 以保证细胞发生重建;②分化后成熟的脂肪细胞内有大量脂滴形成,线粒体的数量和功能对于脂滴及甘油三酯的合成有重要作用。Mitotracker Red 探针对于线粒体的染色是亲脂的,不依赖于线粒体膜电位[11-12]。通过Mitotracker Red 探针标记线粒体(红色)和LC3 免疫荧光标记自噬体(绿色)不仅可以分别观察自噬体和线粒体[13-14],也可以在共聚焦显微镜下观察二者的共定位,以确定线粒体自噬的发生[15-17]。本研究中Mitotracker Red 探针标记检测结果显示:p62 基因缺失细胞中LC3 的点状化和颗粒化增多,并观察到与线粒体的共定位现象增加,表明二者的复合体增多,提示p62 基因缺失在hADSCs 成脂中促进线粒体自噬。本研究使用CsA抑制线粒体自噬,当p62 基因缺失引起线粒体自噬的增加被CsA 抑制后(CsA+p62 基因缺失),其促进成脂的作用也被抑制,表明p62 基因缺失导致的成脂增加与线粒体自噬作用增强有关。关于p62 基因缺失如何会增强线粒体自噬,可能与p62 基因在机体中对线粒体作用的双面性有关:p62 基因既可作为中介分子介导线粒体自噬发生,也可稳定线粒体的形态使其免受损伤而抑制线粒体自噬的发生[18-20]。
综上所述,在hADSCs 脂肪分化过程中,p62基因缺失不仅导致自噬流增加,而且使得自噬流的分布不均:线粒体导向的自噬流增多,保证了hADSCs 分化中所需大量线粒体的更新,使分化进一步延续,脂质合成逐渐增加,最终使得成脂增加。
猜你喜欢
家畜生态学报(2022年2期)2022-02-28 01:52:28
海洋通报(2021年1期)2021-07-23 01:55:14
生物学通报(2021年4期)2021-03-16 05:41:26
复旦学报(医学版)(2019年4期)2019-08-07 04:55:38
中成药(2017年10期)2017-11-16 00:49:52
罕少疾病杂志(2016年2期)2017-01-11 12:27:34
华南农业大学学报(2016年5期)2016-08-05 05:35:04
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:05
河北大学学报(自然科学版)(2015年1期)2015-02-27 13:06:10
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
