
条石鲷精子诱导大黄鱼减数分裂雌核发育
2020-10-21 02:13陈睿毅徐冬冬张琴星孟祥磊
浙江海洋大学学报(自然科学版) 2020年2期
陈睿毅,徐冬冬,楼 宝,张琴星,孟祥磊
(1.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江省海水增养殖重点实验室,浙江舟山 316021;2.三门县农业农村局,浙江三门 317100;3.嵊泗县海洋与渔业局,浙江舟山 202450)
大黄鱼Larimichthys crocea,隶属鲈形目,石首鱼科,黄鱼属。其肉质细嫩、味道鲜美,是我国重要的海水经济鱼类,据不完全统计[1],2018 年大黄鱼养殖产量达20.35 万t,是我国8 大优势出口水产品之一,是一条产业价值百亿的特色鱼。与半滑舌鳎Cynoglossus semilaevis[2]和黄姑鱼Nibea albiflora[3]等类似,大黄鱼也存在着雌性生长快于雄性的现象[4]。因此,通过选育手段实现大黄鱼全雌化养殖,可以缩短大黄鱼养殖周期,提高经济效益。
人工雌核发育技术作为一种有效的性别控制方法已广泛应用于多种鱼类的选育[5-7]。一般来说,雌核发育诱导中用于刺激卵子分裂的“激活源”主要分为“同源精子”和“异源精子”两种。目前,在牙鲆Paralichthys olivaceus 雌核发育的研究中发现,利用同源精子诱导减数分裂雌核发育,获得的子代中有1 尾牙鲆中存在父方基因表达的现象[8],而利用真鲷Pagrus major 精子诱导的雌核发育[9],RAPD 分析结果显示,真鲷精子的遗传信息未在雌核发育二倍体的电泳图中检出。同源精子诱导雌核发育,由于不能保证精子被百分之百的灭活,存在精子遗传物质残留于雌核发育胚胎中的可能性,还存在雌核发育后代中混入正常二倍体的现象。而利用亲缘关系较远的异源精子诱导雌核发育则可有效避免精子灭活不彻底形成正常受精的可能性,存活下来的即为雌核发育二倍体,这在一定程度上也表明异源精子在诱导雌核发育方面拥有更高的准确率。大黄鱼同源精子诱导雌核发育的研究在国内已有报道,吴清明等[10]采用静水压抑制第一次卵裂的方法成功获得了双单倍体大黄鱼,4 个微卫星检测表明6.67%的子代含有父本特有等位基因。苗亮等[11]和汪倩凤等[12]则分别利用未经灭活的鮸鱼精子Miichthys miiuy 和灭活的黄姑鱼精子诱导了大黄鱼减数分裂雌核发育,结果表明两者均能成功激活卵子并发育成仔鱼。
本研究在参考前人相关研究的基础上,利用与大黄鱼亲缘关系更远的条石鲷Oplegnathus fasciatus 精子作为激活源,优化了条石鲷精子激活大黄鱼卵子及诱导其雌核发育的相关参数,并用流式细胞仪检测了雌核发育群体的二倍性,用组织切片的方法证明了雌核发育大黄鱼的全雌性。
1 材料与方法
1.1 精卵的获得
实验用大黄鱼亲鱼来自台州市大陈岛养殖有限公司,2017 年4 月初从大陈岛网箱养殖基地运输至舟山,在室内进行营养强化4 周,水温升至20 ℃后开始实验。从亲鱼群体中(511.74±76.22 g)挑选体格健壮,性腺发育良好的雌性大黄鱼注射促黄体素释放激素A3(LHRH-A3),剂量为8.0~10.0 μg·kg-1,30~36 h 后轻轻挤压大黄鱼的腹部获得卵子,置于烧杯中。实验用条石鲷雄鱼为来自浙江省海洋水产研究所试验场培育的3 龄亲体(323.12±26.32 g),条石鲷雄鱼无需催产,轻轻挤压雄鱼腹部即可获得精子。采集的精子和卵子常温下避光保存备用。
1.2 精子的紫外线灭活
精子灭活所用紫外灯箱中有2 支15 W 的紫外灯(Philips,G15T8),灭活前提前预热15 min。将条石鲷精子用常温Ringer 氏液稀释40 倍,取稀释后的精液5 mL 置于直径9 cm 的玻璃培养皿中,然后将培养皿置于距灯管垂直距离20 cm 处进行灭活。采用紫外辐照计(UV-B(254),北京师范大学光电仪器厂)测量此处辐射强度为1 330~1 350 μW·cm-2。为了筛选最佳的照射剂量,分别设定紫外时间为0 s、15 s、30 s、45 s、60 s、75 s、90 s、120 s、180 s 和240 s。将不同照射时间的精液分别取200 μL 与1 mL 的大黄鱼卵子混合,常温海水激活后置于1 L 烧杯中孵化,孵化温度为21~22 ℃。以上处理均做3 个平行实验,统计其孵化率,得到最佳精子灭活时间。
1.3 人工授精与冷休克
参考许建和等[13]同源精子诱导大黄鱼雌核发育的方法,略作修改,具体步骤为:利用灭活后的条石鲷精子对大黄鱼卵子进行人工授精,授精2 min 后将受精卵置于3 ℃海水中处理10 min,然后21~22 ℃水温下孵化。另设单倍体对照组(灭活精子授精,未经冷休克)和正常对照组(鲜精授精),实验组和对照组均做了3 个平行家系。
1.4 胚胎发育观察及倍性检测
在受精卵孵化期间,采用NIKON-MSZ800 解剖镜观察胚胎发育并拍照。仔鱼出膜后,随机挑选单倍体、正常二倍体和雌核发育二倍体各30 尾,采用流式细胞仪Ploidy Analyzer (BD FACSCalibur,American)进行分析测定。
1.5 雌核发育二倍体的性腺切片观察
雌核发育大黄鱼4 月龄时,随机挑选30 尾测量其体长和体质量,采用麻醉剂MS222 麻醉后解剖取性腺,固定于波恩液中;然后将样品进行石蜡包埋、组织切片和苏木素-伊红染色。采用Olympus MS800 显微镜对组织切片进行观察,统计性别比例。
1.6 数据统计分析
本实验中受精率和孵化率的统计方法为:受精率为发育至原肠期的受精卵数与总授精卵数之比;孵化率为孵化个体数与原肠期正常胚胎数之比。用SPSS19.0 进行单因素方差分析和Duncan 氏多重比较。
2 结果与分析
2.1 紫外线灭活
条石鲷精子在不同紫外线强度灭活后,经其授精的大黄鱼卵子表现出不同的孵化率(图1)。从图1 可知,未经灭活的条石鲷精子其孵化率为63.12%±2.78%;当其在紫外线下照射5 s 之后,孵化率降至33.18%±2.34%;25 s 时孵化率达到最低值为18.58%±3.23%。之后随着灭活时间的延长其孵化率逐渐升高,至60 s 时最高,为47.39%±2.24%;60 s之后孵化率又逐渐降低,呈现出显著的Hertwig 效应。结果表明,条石鲷精子紫外线灭活的最佳时间为60 s,此时累积辐射量为80 mJ·cm-2。
2.2 受精率和孵化率
大黄鱼正常对照组的受精率和孵化率分别为67.32%±8.39%和73.18%±4.21%,显著高于实验组的39.36%±3.47%和34.28%±3.12%(P<0.01)(表1)。

图1 紫外线照射时间对大黄鱼孵化率的影响Fig.1 The influence of UV irradiation duration on the hatching rate of L.crocea

表1 对照组和实验组的受精率和孵化率Tab.1 The fertilization rate and incubation rate of control and experimental group
2.3 胚胎发育观察及倍性检测
未经冷休克处理的受精卵孵化后均为畸形,表现出显著的“单倍体综合征”,且出膜后全部死亡(图2)。倍性检测结果显示,雌核发育二倍体和正常二倍体的DNA 相对含量均为200,而单倍体的DNA 含量为100(图2)。说明条石鲷精子中的DNA 未进入雌核发育大黄鱼的基因组;同时还表明通过冷休克的方式可以实现使大黄鱼染色体数目加倍的目的。
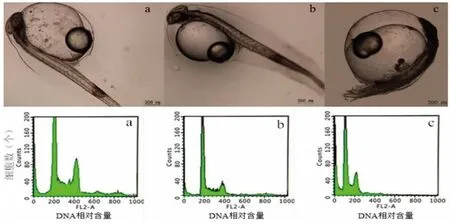
图2 大黄鱼雌核发育二倍体(a)、正常二倍体(b)和单倍体(c)(人工授精50 h 后,孵化水温21 ℃)Fig.2 External morphologies and patterns of gynogenetic diploid,diploid control and haploid larvae of L.crocea (50 h,T=21 ℃)
2.4 性腺组织切片检测
4 月龄时大黄鱼平均体长7.88±0.57 cm,平均体质量为8.32±0.63 g,30 尾雌核发育大黄鱼性腺切片均能观察到初级卵母细胞和卵巢腔,表明雌核发育大黄鱼性腺发育为卵巢。
3 讨论

图3 4 月龄雌核发育大黄鱼性腺组织切片Fig.3 Gonadal histology of gynogenetic large yellow croaker at 4 months of age
人工雌核发育可分为减数分裂雌核发育和有丝分裂雌核发育,前者通过抑制第二极体排出,后者则通过抑制第一次有丝分裂而使染色体加倍。但两种方法均需要利用精子对卵子进行激活,因此“激活源”的选择对于雌核发育的成功率至关重要。有学者指出[15],大多数属间以上的杂交相容性都很低,大部分胚胎在孵化期前后陆续死亡,表现出极低的孵化率。条石鲷和大黄鱼分属不同科,相比于黄姑鱼和鮸鱼[11-12],在遗传进化上条石鲷与大黄鱼亲缘关系更远。实验中发现,条石鲷和大黄鱼的杂种虽然可以出膜,但在开口之前全部死亡,这样即使有少量雄核的遗传物质未被完全灭活,个体也会在出膜后自然淘汰,避免了正常二倍体混入的可能。
精子的遗传物质的失活是影响雌核发育能否成功的关键因素[16],但对于诱导雌核发育究竟是选择同源精子还是异源精子,目前尚无统一结论。有研究表明:同源精子成功诱导雌核发育所需紫外辐射剂量更高[12],而使用异源精子诱导雌核发育能显著提高雌核发育子代成活率[17-18]。本研究中,将条石鲷精子用常温Ringer 氏液稀释40 倍后,置于紫外灯下进行灭活,当辐射强度为80 mJ·cm-2时,大黄鱼受精率和孵化率最高。而汪倩凤等[12]利用黄姑鱼精子诱导大黄鱼雌核发育,黄姑鱼精子紫外灭活处理的最适剂量则为198 mJ·cm-2,另有研究表明[19]同源精子诱导大黄鱼雌核发育,精子紫外灭活剂量在253~406 mJ·cm-2均能成功诱导,这些实验结果与本研究存在较大差异,可能是因为本研究中精液灭活是在常温下进行,而上述其他研究则是在4 ℃下或冰面上进行。精子的适宜紫外灭活剂量随鱼种类不同而存在较大差异,如条斑星鲽Verasper moseri 精子为40~45 mJ·cm-2[20],大西洋鳕Gadus morhua 精子为221 mJ·cm-2[21],斜带石斑鱼Epinephelus coioides 精子为518 mJ·cm-2[22]。同样,精子保存液的种类也会对同种鱼类精子的灭活剂量产生影响,如黄姑鱼精子用不同的精子保存液稀释后(Ringer 保存液和Hanks 保存液),其最适紫外灭活剂量分别为420 mJ·cm-2[23]和228 mJ·cm-2[24],存在较大差异。此外,灭活时精液的温度、精液的稀释倍数和精液厚度等因素[25],也会造成最适灭活剂量的差异。因此实际操作中,一方面可以通过显微镜观察紫外线照射后精子的活力,另一方面还可以通过观察雌核发育后代的孵化率来判断最适的灭活剂量。
雌核发育的目的一般是为了获得纯系的单性别群体,但由于人为处理并不能使精子100%灭活,因而必须有充分的证据证明精子在雌核发育过程中没有提供遗传物质。本研究中,利用亲缘关系较远的条石鲷精子诱导大黄鱼雌核发育,有效避免了精子灭活不彻底形成正常受精卵的可能性,存活下来的个体经倍性检测证实为二倍体,性腺组织切片也证明了它们的全雌性,这表明条石鲷精子可以成功诱导大黄鱼雌核发育,是一种合适的“激活源”。鱼类的性别决定机制多样而复杂,雌核发育可以作为一种重要的手段用以判别鱼类的性别决定机制[26-27]。雌核发育个体所有遗传物质均来自于母本,理论上,XX/XY 性别决定机制的个体雌核发育后代全部为XX 个体,即雌性个体。本研究中,利用性腺组织切片的方法来鉴定大黄鱼雌核发育群体的性别,结果表明其100%为雌性,这也证明大黄鱼性别为雌性配子同型性别决定型,即为XX/XY 决定型。
综上,本实验研究表明,条石鲷精子是诱导大黄鱼雌核发育的一种合适的激活源,水温3 ℃下,授精2 min 后,冷休克10 min,可以获得大黄鱼减数分裂雌核发育二倍体,并且雌核发育群体具有全雌性,这也从侧面表明大黄鱼的性别决定为XX/XY。
猜你喜欢
海洋开发与管理(2022年10期)2022-11-11
辽河(2022年5期)2022-07-07
农产品市场周刊(2021年17期)2021-09-28
数学大王·中高年级(2020年4期)2020-04-19
当代水产(2019年6期)2019-07-25
大众考古(2017年8期)2017-12-12
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
福建文学(2015年12期)2015-12-07
江苏农业科学(2014年9期)2014-11-15
