
木棉的高抗旱性和其速生性适合修复海南已退化的森林
2020-10-21 08:05罗金环陈广武谭照远
海南大学学报(自然科学版) 2020年3期
罗金环,陈 斌,陈广武,谭照远,张 辉
(1. 三亚林业科学研究院,海南 三亚 572000; 2. 三亚市林业局,海南 三亚 572000;3. 海南大学林学院/海南五指山森林生态系统国家定位观测研究站,海南 海口 570228;4. 海南大学林学院 热带特色林木花卉遗传与种质创新教育部重点实验室,海南 海口 570228)
人类的生产活动(如采矿和农业生产)导致了全世界范围内非常高的森林退化,这已成为对全球生物多样性和生态系统的主要威胁[1-4],基于再造林已经被认为是能够部分缓解森林退化的一种补救方法[5-6],因此,世界各地实施了许多以减轻这些影响为目的的再造林项目[7-11].然而,迄今为止,世界各地多以单一种植外来速生树种和商用树种作为主要的再造林方式,并依赖这种方式来修复已退化的森林生态系统[12].
虽然单一种植这些外来树种可以达到迅速修复已退化森林的目的,但它们也可能增加后期维持土壤水分和养分的成本,并具有非常高的森林病虫害暴发的风险[13-14].因此,许多研究强调,修复已退化森林生态系统的最关键步骤是要使用合适的本地树种,而不是外来树种[15-16].然而,选择合适的本地树种要比选择外来树种复杂得多,也更具挑战性[17-18].
目前所开发的基于性状的方法可以为选择合适的本地树种提供良好的途径[19-20].该方法认为,植物的功能性状能直接反映植物对不同的非生物和生物环境的适应性[21-23].因此,利用功能性状可寻找出既具有较高的存活率和生长率,又能很好地适应高度退化的生态系统环境的本地树种,并以其来修复已退化森林的生态系统.
木棉是一种广泛分布于中国海南岛的本地热带树种[24],也是一种可用于雕塑的需求量极大的树种,因此其幼苗已被广泛种植并具有极佳的经济效益[25].此外,由于木棉还可以很好地适应恶劣的环境(如干旱胁迫)[26],因此,它可能会具有很高的可用于修复海南已退化热带森林生态系统的潜力.然而,几乎没有研究证明它是否真的适合在海南岛修复已退化的热带森林生态系统.鉴此,2016年,在中国海南省三亚市,当地政府在抱坡岭上单一种植了木棉,并以其来恢复抱坡岭0.2 km2范围内高度退化的热带森林,这些用于修复抱坡岭退化森林的木棉将有助于检验木棉是否是适合于修复海南已退化热带森林的本地树种.由于海南岛存在季节性的干旱胁迫[27],如果这些用于修复抱坡岭已退化森林的木棉在旱季既具有高抗旱性和速生性,又具有很高的存活率,那么木棉将会是适合修复海南已退化森林的树种.基于此,本研究记录了其在抱坡岭的存活率,同时于旱季(1月份),比较了抱坡岭的木棉和苗圃培育的木棉在功能性状方面的差异,进而检验木棉在旱季是否确实具有较高的存活率、生长率和抗旱性,并以此来确定木棉是否可以作为修复海南已退化热带森林的良好候选本地树种.
1 材料与方法
1.1 研究地点概况研究区位于中国海南省三亚市的抱坡岭(109°51′01′′E, 18°31′99′′N),该岭是石灰岩山,属于热带季风海洋性气候,年平均温度为28 ℃.年平均降雨量为1 500 mm,其中6~10月降雨量最多(91%)[28-29],其典型的植被为物种丰富的热带季风阔叶林.为了生产水泥,石灰岩山被不间断地开采了20年,使抱坡岭的一部分(0.2 km2的面积)已经成为一个高度退化的裸露岩石山,不能种植任何植物.2016年,三亚市政府在这片0.2 km2的区域实施了单一种植木棉幼苗的再造林工程,种植密度保持在每公顷80~100棵,我们记录了2016—2019年木棉的存活率:

1.2 取样方法由于旱季的降雨量仅为雨季的1/10,所以本研究只关注木棉在旱季的响应.我们于2019年1月(旱季)在抱坡岭,从5个胸径相当的木棉个体中采集了20片完全展开、健康且暴露在阳光下的叶片;同时也在最初用于抱坡岭木棉造林工程的木棉幼苗的苗圃中使用相同的取样方法,收集了20片充分展开的、健康的、暴露在阳光下的木棉叶片.苗圃的木棉是2016年培育且准备用于修复抱坡岭的,但是没有使用完,所以苗圃的木棉和抱坡岭的木棉除了生长环境不一样之外没有任何差异.因此对比抱坡岭木棉和苗圃木棉的差异,可以得知木棉在抱坡岭的生长过程中是否已具有高抗旱性和快速生长的特性.
1.3 性状测量方法我们利用从抱坡岭和苗圃采集的叶片样本来测量与快速生长和抗旱性高度相关的功能性状[上表皮厚度(μm)、栅栏组织厚度(μm)、海绵组织厚度(μm)、下表皮厚度(μm)、气孔密度(数量·mm-2)、蒸腾速率(mol·m-2·s-1)、最大光合速率(mol·m-2·s-1)、气孔导度(mmol·m-2·s-1)和叶片膨压丧失点(MPa)].这些功能特征的详细测量步骤如下所示.
1.3.1 叶片解剖性状的测定于抱坡岭和苗圃中分别从5株独立的木棉幼树上采集20片成熟的、暴露在阳光下的叶片,然后将所有叶片样品放在乙醇系列溶液(50%、70%、85%、95%和100%)中逐渐脱水,脱水后用热石蜡浸润,然后用徕卡DM2500光学显微镜(Leica Microsystems Vertrieb GmbH, Wetzlar, Germany)测量叶片、上表皮、下表皮、海绵组织和栅栏组织的厚度,并用剃须刀片切割远轴表皮以测量气孔密度.最后,将所有切片均置于载玻片上,在徕卡DM2500显微镜下观察.
1.3.2 最大光合速率、蒸腾速率和气孔导度的测量在晴天的上午9点至11点,利用Li-6400便携式光合系统仪(Li-6400, Li-Cor, Lincoln, Nebraska, USA)测量木棉幼树的最大光合速率、蒸腾速率和气孔导度.基于初步试验,光照强度设定在1 500 μmol·m-2·s-1,以确保测量出所有树的光饱和速率;二氧化碳设定为390 μmol·mol-1,气温设定为28 ℃.在记录数据之前,将叶片暴露在上述条件下约5 min,使光合参数稳定下来.
1.3.3 叶片膨压丧失点的测定于抱坡岭和苗圃中,分别从5株独立的木棉幼树上采集带叶的枝干,采集完毕后立即送至实验室.在实验室里,将枝干的基部浸泡在蒸馏水中后,重新剪切枝干的基部,然后复水至叶片水势大于-0.05 MPa,接着,称量叶片的重量以获得初始鲜重,再立即将叶片置于压力室中以确定初始水势,在叶片摘下之后,定期反复测量叶片的鲜重和水势,直至水势基本稳定为止.最后,将叶片在70 ℃下烘干72 h,测量其干重.而叶片膨压损失点则采用Schulte 和Hinckley发明的压力-体积关系分析模型来进行测定[30].
1.4 数据处理我们首先利用 Wilcoxon符号秩检验(Wilcoxon rank sum tests)定量探究抱坡岭旱季(1月)和雨季(7月)的降雨量是否存在显著差异;同时也采用 Wilcoxon 符号秩检验来定量分析抱坡岭木棉和苗圃木棉的9个与生长和抗旱高度相关的功能性状(上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度、气孔密度、最大光合速率、蒸腾速率、气孔导度和叶片膨压丧失点),并据此来判断它们之间是否存在显著差异.最后,利用主成分分析(PCA)来评比这9个性状中哪些性状能显著区别抱坡岭木棉和苗圃木棉,以上的数据处理均在软件R 3.6.1中完成.
2 结果与分析
2.1 抱坡岭旱季与雨季的平均降雨量基于三亚气象局近三年的降雨数据得知,抱坡岭旱季(1月)的平均降水量仅为雨季降雨量(7月)的1/10(图1),显著低于雨季的平均降水(Wilcoxon 符号秩检验P<0.05, 表1),因此生长在抱坡岭的木棉在旱季有可能会遭受干旱胁迫.
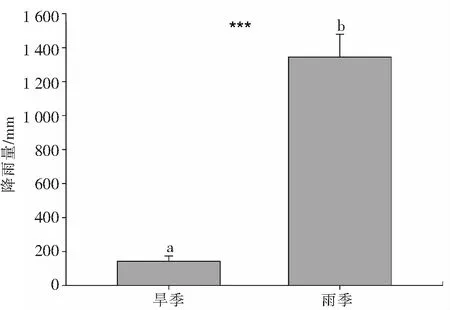
a、b表示存在显著差异(P<0.05),基于Wilcoxon 符号秩检验得出

表1 基于Wilcoxon 符号秩检验的抱坡岭旱季(1月)和湿季(7月)平均降雨量的差异对比
2.2 抱坡岭木棉的存活率和其与苗圃木棉在旱季的功能性状差异在抱坡岭,木棉幼树的存活率非常高(92%±4%),由图2可以看出,在旱季,抱坡岭木棉与生长相关的功能性状(上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度、蒸腾速率、最大光合速率)均显著高于苗圃木棉与生长相关的功能性状(图3,Wilcoxon 符号秩检验P<0.05, 表2),其中,叶片的上表皮厚度和下表皮厚度是苗圃木棉的2倍左右,另外,在两个解剖性状方面——栅栏组织厚度和海绵组织厚度,抱坡岭木棉是苗圃木棉的2倍左右,抱坡岭木棉的蒸腾速率大约是苗圃木棉的5倍,抱坡岭木棉的最大光合速率大约是苗圃木棉的3倍;但是抱坡岭木棉叶片的气孔密度却是苗圃木棉的1/3~1/2,抱坡岭木棉的气孔导度大约是苗圃木棉的1/6,抱坡岭木棉与苗圃木棉的叶片膨压丧失点都为负值,并且抱坡岭木棉的叶片膨压丧失点为苗圃木棉的3~4倍.
对于所测量的与生长速率相关的6个功能性状,均是抱坡岭木棉显著高于苗圃木棉,而对于气孔密度、气孔导度和叶片膨压丧失点,则是抱坡岭木棉显著低于苗圃木棉(图2,Wilcoxon 符号秩检验P<0.05, 表2).
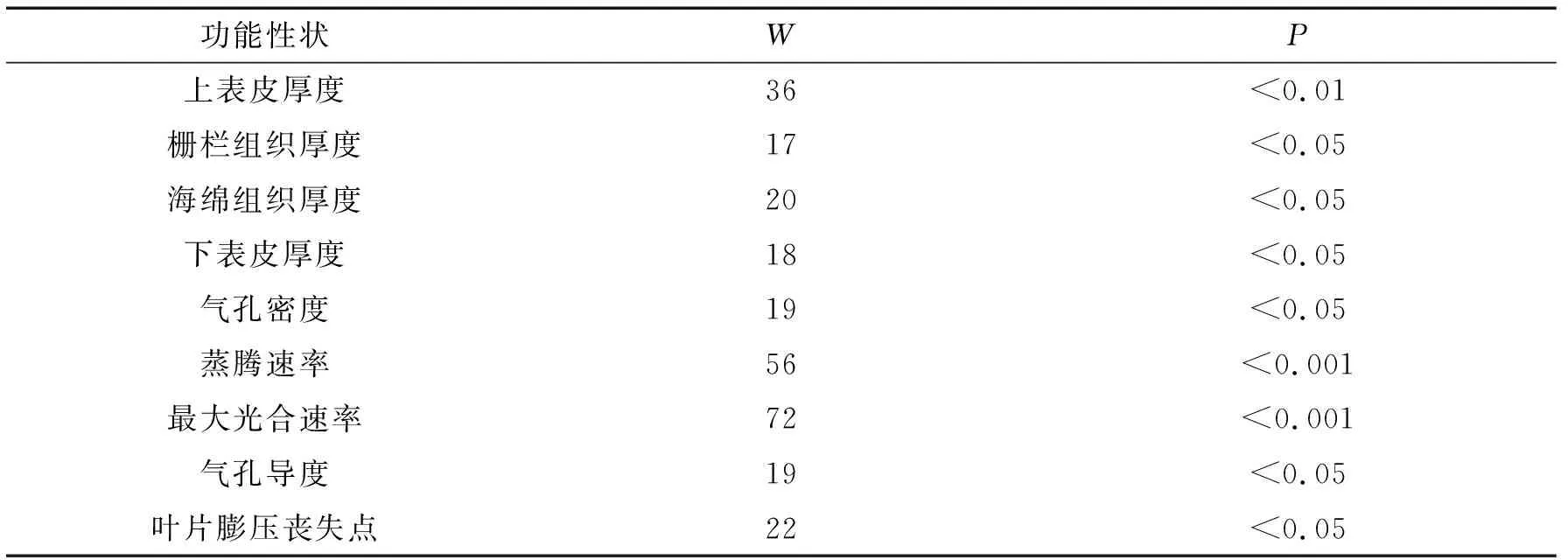
表2 基于Wilcoxon 符号秩检验的抱坡岭木棉和苗圃木棉的功能性状差异
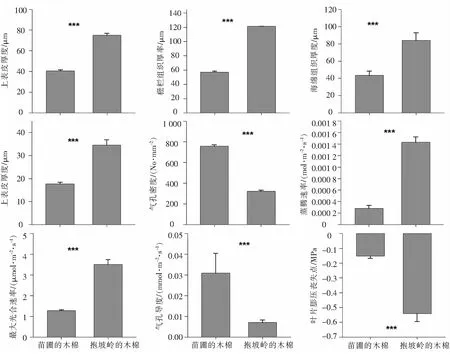
***表示存在显著差异(P<0.05),基于Wilcoxon 符号秩检验得出
2.3 抱坡岭木棉和苗圃木棉的主成分分析表3和图3为主成分分析的结果,PC1的贡献率高达92.19%,PC2的贡献率是4.18%,故取主成分分析的前两轴,且结果表明,只有气孔导度最能区分抱坡岭木棉和苗圃木棉 (PC1和PC2>0.5, 表3),而其他8个功能性状并不能很好地区分抱坡岭木棉和苗圃木棉(PC1和PC2<0.5, 表3).

表3 基于9个植物功能性状得到的抱坡岭木棉和苗圃木棉主成分分析的前两个轴

图3 抱坡岭木棉和苗圃木棉9个功能性状的主成分分析图
3 讨 论
通过性状对比分析发现,抱坡岭木棉的光合速率显著比苗圃木棉的高,大约是苗圃木棉的3倍.光合速率能影响植物的能量平衡,通常,较高的光合速率与较快的生长速率相联系[31-32],这说明木棉具有较高的生长速率.由于植物功能是由其相对应的组织结构来支撑,因此,更高的光合速率会需要更好的叶片解剖性状(例如上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度和气孔密度)来支撑[33-34].这就是为什么在抱坡岭中观察到的木棉相较于苗圃中的木棉显著具有更厚的上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度和气孔密度的原因.同时,由于光合速率和蒸腾速率是显著耦合的[35],这也就是为什么抱坡岭木棉比苗圃木棉具有更高蒸腾速率的原因了.这些结果表明,木棉具有与较高生长速率相适宜的功能性状.
研究结果清楚地显示,抱坡岭在旱季的平均降雨量仅为雨季的1/10,说明抱坡岭木棉有很高的遭受干旱胁迫的风险.通过对比抱坡岭木棉叶片和苗圃木棉叶片的膨压丧失点差异,我们发现,抱坡岭木棉的叶片膨压丧失点要比苗圃的低得多.当叶片不能获得充足的水分供应时,通常会出现叶片膨压丧失点偏低的情况[36],因此抱坡岭木棉的确遭受了干旱胁迫.较低的叶片膨压丧失点也能表征较高的植物抗旱性[36],因此抱坡岭的木棉在旱季具有高抗旱性.
然而值得注意的是,高光合速率应该导致较高的气孔密度和气孔导度[37-38],然而我们发现,与苗圃木棉相比,抱坡岭木棉的气孔导度要低得多.较低的气孔导度和气孔密度与气孔关闭高度相关,其可导致叶片膨压丧失点降低以适应干旱胁迫[39-41].抱坡岭木棉的叶片膨压丧失点也的确要比苗圃的低得多,因此,抱坡岭木棉在旱季可通过气孔关闭来降低叶片膨压丧失点,从而使其具有高抗旱性以适应干旱胁迫.主成分分析结果显示,仅有气孔导度能显著区分抱坡岭木棉与苗圃木棉.因此,在旱季,较低的气孔导度在一定程度上会导致抱坡岭木棉的气孔关闭,从而使其能保持较低的叶片膨压丧失点和具有较高的抗旱性以及较高的生长速率[36].
4 结 论
综上所述,在抱坡岭种植木棉以修复高度退化的热带森林时,往往会引发木棉叶片的气孔关闭,使其具有较高的抗旱性,进而使其在旱季避免受到干旱胁迫的影响.此外,抱坡岭木棉还具有与较高的光合速率相适应的功能性状,这使其具有了较高的生长速率,从而使其存活率可达到很高的水平(92%±4%).因此,木棉的高抗旱性、速生性和高存活率使得其成为一个非常适合修复海南已退化热带森林的本地树种.
猜你喜欢
生态学报(2022年18期)2022-10-13
作物研究(2022年4期)2022-09-21
东北林业大学学报(2022年4期)2022-05-16
农业工程学报(2022年3期)2022-04-16
农业工程学报(2021年3期)2021-04-15
歌海(2021年6期)2021-02-01
幼儿园(2020年15期)2020-11-24
艺术家(2020年5期)2020-07-09
海峡姐妹(2019年5期)2019-06-18
江苏农业科学(2017年1期)2017-02-27
