
柠檬桉3个不同种源光响应特征分析
2020-10-10 03:08:16谢春俊梁丽华韦大勇唐庆兰
桉树科技 2020年3期
谢春俊,梁丽华,韦大勇,唐庆兰
(1.广西国有三门江林场,广西 柳州 545000;2.广西国有东门林场,广西 崇左 532108;3.广西国有维都林场,广西 来宾 546100;4.广西壮族自治区林业科学研究院,广西 南宁 530002)
柠檬桉(Corymbia citriodora)是桃金娘科(Myrtacese)伞房属(Corymbia)树种,分布在澳大利亚昆士兰州中部和北部的沿海区域,地跨热带和亚热带(15.5 ~ 25.0°S)。1 ~ 8 a生胸径和高生长较快,10 a后材积增长较快,18 ~ 25 a达数量成熟。木材边材呈白色,心材为深棕色,木材纹理直或偶尔呈波纹和交错状,材质粗糙、坚硬、结实、有耐性、易加工。成熟材的基本密度可达950 ~ 1 010 kg·m-3,可作桥梁、建筑、地板等用材[1],还可用于造纸、蒸馏精油等[2]。
柠檬桉于1915年首次引种至我国,以路旁树为主。1954年,广东粤西桉树林场和广西合浦县山口国营林场开始经营柠檬桉。50年代末,广西、云南、浙江等地进行柠檬桉引种试验。中澳技术合作东门桉树示范林项目(1982—1989)引种了多个桉树树种,其中柠檬桉幼龄材纤维长、易漂白、纸浆得率高、质量好,是较好的纸浆材。目前国内柠檬桉的研究主要集中在柠檬桉叶片和嫩枝的挥发性油类的提取、成分分析等[3-7],对柠檬桉的引种、选育研究不多[8-9]。本文对3个种源柠檬桉的光响应特征参数进行分析,以期为引种和栽培提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于广西南宁市郊广西林业科学研究院科研试验苗圃基地(22°56′N,108°56′E),海拔约80 m,≥ 10℃积温为7 200℃,年平均气温21.7℃,极端最低温-1.5℃,极端最高温39.4℃,全年无霜期达360 d,年均降雨量为1 300 mm,年均相对湿度约为80%。
1.2 参试种源
2013年12月从澳大利亚联邦科学与工业研究组织引进柠檬桉3个种源(表1)。

表1 柠檬桉参试种源来源地概况
1.3 光合测定
2016年1月播种,当苗木长至30 cm高后,将其移至直径30 cm的塑料杯内,当年10月份进行光合测定。
用美国LI-COR公司生产的LI-COR6400光合作用仪进行测定,测定时间为晴天上午9:00―11:00。光响应曲线的测定:每种源选取3株平均样木,每株选取从树枝顶部往下第一片向阳的功能叶进行光响应曲线的测定,光强(PAR)梯度分别设定为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、500、200、100、50、和0 μmol·m-2·s-1。每个处理重复3次。
1.4 数据处理及分析
分别采用SPSS 19.0和EXCEL 2007进行数据分析和作图。
2 结果与分析
2.1 不同种源柠檬桉光响应曲线
不同种源柠檬桉净光合速率Pn随光强变化的曲线见图1。由图1可知,3个种源的柠檬桉光响应曲线变化趋势一致,其净光合速率均随光强增加而升高。光强低于1 000 μmol·m-2·s-1时,3个种源的曲线几乎重叠;光强大于1 000 μmol·m-2·s-1后,种源19961的净光合速率高于种源20014和19693,种源19961的Pn最大值为24.719 3 μmol CO2·m-2·s-1,种源20014和19693的Pn最大值分别为22.560 2、22.681 8 μmol CO2·m-2·s-1,2个种源相差不大。

图1 不同种源柠檬桉光响应曲线
不同种源的光响应曲线的二次回归拟合和光强200 μmol·m-2·s-1以下的线性拟合(表 2),可计算出一系列光响应特征参数(表3)。由表2可知,曲线拟合相关系数R2≥0.946,说明二项式回归方程可较好反映柠檬桉净光合速率随光强的变化情况。3个柠檬桉种源的光饱和点较高,种源间相差不大,在1 495~ 1 585 μmol·m-2·s-1之间,利用强光的能力较强。光补偿点差异明显,种源 20014的光补偿点最高(52.59 μmol·m-2·s-1),利用弱光的能力最差,19961次之(33.13 μmol·m-2·s-1),利用弱光的能力中等,19693 最低(18.67 μmol·m-2·s-1),利用弱光的能力较强。
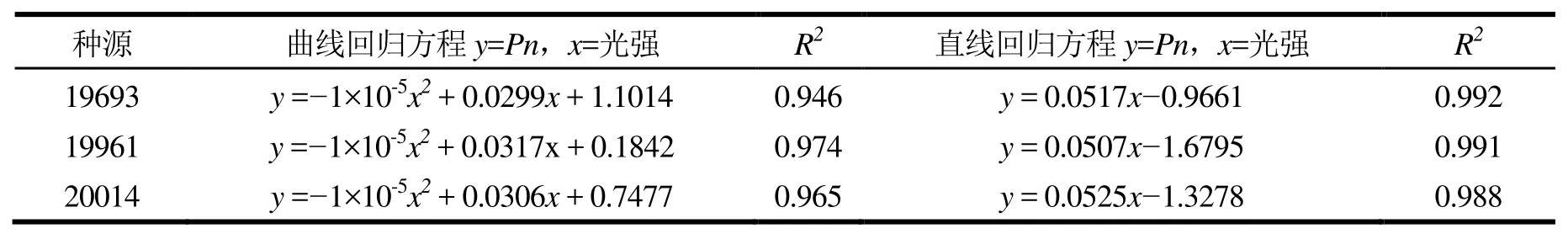
表2 二项式回归方程和光强在200 μmol·m-2·s-1以下的直线回归方程

表3 3个种源柠檬桉光响应特征参数
2.2 不同种源柠檬桉气孔导度Gs响应曲线
由图2可知,3个柠檬桉种源的气孔导度Gs随光强的增强而缓慢上升,各种源变化趋势一致,但种源间变化幅度存在一定的差异。种源19693气孔导度随光梯度的变化幅度最大,变化幅度为0.143~ 0.613 mol H2O·m-2·s-1;种源19661 次之,为0.205~ 0.541 mol H2O·m-2·s-1,种源 20014 较小,0.153 ~0.405 mol H2O·m-2·s-1。
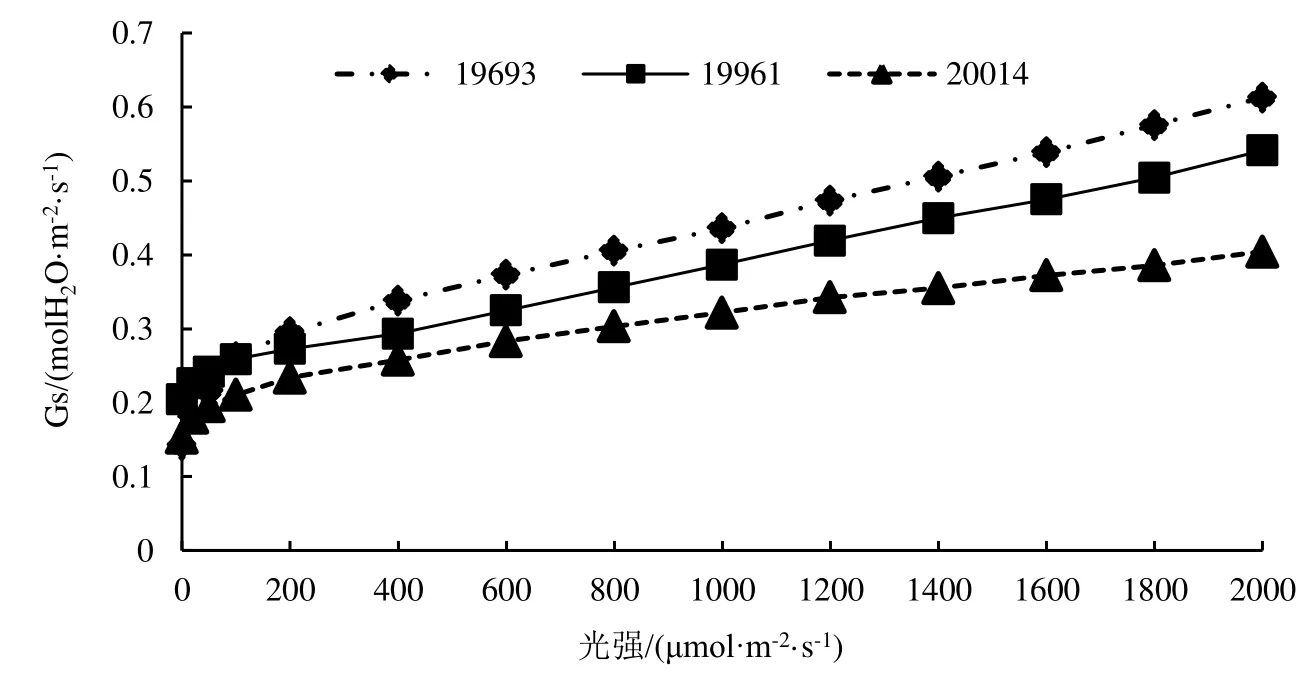
图2 不同种源柠檬桉气孔导度Gs光响应曲线
2.3 不同种源柠檬桉胞间CO2浓度Ci响应曲线
由图3可知,胞间CO2浓度Ci随光强的增大而下降,3个种源的下降趋势基本一致。光强低于400 μmol·m-2·s-1时,各种源胞间 CO2浓度随光强的增加而下降趋势明显,光强在 400 ~ 1 000 μmol·m-2·s-1之间胞间 CO2浓度下降缓慢,光强大于1 200 μmol·m-2·s-1时,各种源的胞间 CO2浓度缓慢上升,上升幅度比较小。各种源间胞间CO2浓度存在一定的差别,种源19693各光梯度的胞间CO2浓度均比另外两个种源的大,种源19961和20014相差不大。
2.4 不同种源柠檬桉蒸腾速率 Tr和水分利用效率WUE的响应曲线
由图 4可知,不同种源的柠檬桉蒸腾速率 Tr随光强增强而增大,各种源曲线变化趋势一致,与气孔导度随光强变化的趋势相似。种源19961的蒸腾速率最大,为4.468 ~ 8.252 mmol H2O·m-2·s-1;种源19693和20014变化范围相近,在1.939 ~ 5.606 mmol H2O·m-2·s-1之间。
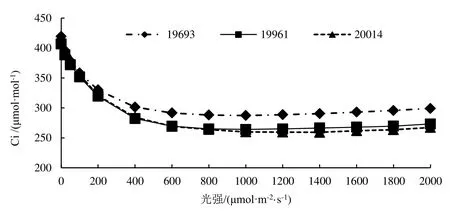
图3 不同种源柠檬桉胞间CO2浓度Ci光响应曲线

图4 不同种源柠檬桉蒸腾速率Tr光响应曲线
柠檬桉各种源水分利用效率WUE随光强增强而增大,各种源变化趋势基本一致,且与净光合速率随光强变化的趋势相似(图5)。各种源在光强低于400 μmol·m-2·s-1时,水分利用效率迅速增加,在光强为 600 ~ 800 μmol·m-2·s-1时,水分利用效率达最大值,随后水分利用效率随光强的增加而趋向平缓,甚至下降。各种源间水分利用效率存在一定的差异,种源19693和20014变化幅度相差不大,在-0.746 ~4.251 μmol·mol-1之间;种源 19961变化较小,在-0.489 ~ 2.995 μmol·mol-1之间。

图5 不同种源柠檬桉水分利用效率WUE光响应曲线
3 结论
(1) 种源19961的Pn最大值为24.719 3 μmol CO2·m-2·s-1,种源 20014 和 19693 的Pn最大值分别为 22.560 2、22.681 8 μmol CO2·m-2·s-1。
(2) 3个不同柠檬桉种源的光补偿点在1 495 ~1 585 μmol·m-2·s-1之间,表现出明显的阳生树种的光强利用特征,对强光的利用能力较强,与巨桉(E.grandis)、尾叶桉(E.urophylla)、尾巨桉(E.urophylla×E. grandis)、巨尾桉(E. grandis×E.urophylla)、斑皮柠檬桉(C.citriodorassp.variegata)和斑皮桉(E.maculata)等树种的光饱和点相似[10-13]。种源 19693的光补偿点最低,为 18.67μmol·m-2·s-1,对弱光的利用能力最强,种源20014光补偿点最高,为52.59 μmol·m-2·s-1,利用弱光的能力弱。
(3) 植物叶片的净光合速率Pn和水分利用效率WUE与气孔导度Gs密切相关。气孔导度是反映植物对环境适应性的种源参数之一。不同种源柠檬桉的 Gs随光强增加而增大,无气孔关闭的情形,说明各种源对不同环境表现出较好的适应性。气孔是植物吸收二氧化碳、水分蒸腾的重要通道,净光合速率Pn和蒸腾速率Tr的比值为植物叶片的水分利用效率 WUE,是用来评价植物生长适应性的重要生态指标之一。结果表明,种源19693和20014的WUE较种源19961的高,说明种源19961需要水分丰富的环境条件才能发挥其生长优势。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
种子(2023年7期)2023-09-24 07:40:54
特产研究(2022年6期)2023-01-17 05:05:02
湖北农业科学(2021年13期)2021-08-04 03:56:18
新疆农业科技(2021年2期)2021-07-03 06:39:34
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
辽宁林业科技(2021年1期)2021-03-16 09:17:08
节水灌溉(2020年9期)2020-09-16 00:32:12
中学生物学(2020年12期)2020-03-30 07:27:46
