
花鮈骨、唇鮈骨及其杂交 F1代耗氧率和窒息点的比较研究
2020-10-10 02:28杨壮志李良玉魏文燕张小丽刘家星陶丽竹
中国饲料 2020年17期
陈 健, 杨壮志*, 陈 霞, 杨 淞, 李良玉,魏文燕,杨 马,唐 洪, 张小丽, 刘家星,陶丽竹
(1.成都市农林科学院,四川成都 611130;2.四川农业大学,四川成都 611130)
花鮈骨(Hemibarbus maculatus Bleeker)隶属鲤形目,鲤科,鮈亚科,鮈骨属,俗称大鼓眼、麻鲤、麻沙根,分布较广,自长江以南至黑龙江水系均有分布,是常见的中小型经济鱼类。花鮈骨生长速度较慢,常见个体20~30 g,作为野生鱼受消费者青睐,市场接受度较高 (王志林 等,2003)。唇鮈骨(Hemibarbus labeo pallas)鲤形目,鲤科,鮈亚科,鮈骨属,俗称土风鱼、重唇鱼,在我国各大水系均有分布。唇鮈骨个体较大,生长较快,以动物性饵料为食如水生昆虫的幼虫、摇蚊幼虫以及虾类,也摄取小型软体动物。3龄鱼可达性成熟,生殖季节在3~4月,怀卵量随年龄和个体大小有较大差异。
近年来,课题组为了改良花鮈骨、唇鮈骨生长性状,将花鮈骨(♀)与唇鮈骨(♂)进行杂交,获得了在生长性状和营养品质方面上均具有杂种优势的F1代,在养殖生产中得到了广泛推广,对花鮈骨、唇鮈骨及杂交F1代养殖业的发展起到了积极地推动作用。
耗氧率可直接或间接地反映鱼类的新陈代谢规律、生理状况和代谢水平,耗氧率是研究鱼类新陈代谢规律的重要指标之一(Cao等,2015;Saucedo 等,2003;Kita 等,1996),窒息点则是鱼类耐低氧的极限指标。耗氧率和窒息点数据测定结果可为研究鱼类能量代谢提供基础依据。近年来,国内外学者进行了大量的鱼类呼吸代谢方面的研究,廖志洪等(2004)进行了黄颡鱼仔、稚、幼鱼耗氧率及氨氮排泄率的初步研究,金一春等(2009)进行了罗非鱼耗氧率、窒息点的初步研究,张永泉等(2010)进行了哲罗鱼耗氧率、窒息点的初步研究;但有关花鮈骨、唇鮈骨呼吸代谢方面的研究鲜见报道。本试验以花鮈骨、唇鮈骨及其杂交F1代幼鱼为研究对象,对其耗氧率和临界窒息点进行试验与比较研究,并分析了不同温度下窒息点及昼夜下耗氧率的差别,旨在丰富其呼吸生理学理论,为F1代杂交优势苗种培育、健康养殖及活鱼运输等提供理论依据。
1 材料与方法
1.1 试验材料 试验鱼花鮈骨、唇鮈骨及花鮈骨(♀)×唇鮈骨(♂)F1 代幼鱼(以下称杂交鮈骨)均为蒲江华昱水产养殖场人工繁育。选择规格基本一致、健康无病、生长良好的试验鱼,在试验室孵化桶中停食暂养3 d后用于试验,试验用水为充分曝气的自来水,pH 7.0左右,溶氧含量5 mg/L以上。各组试验鱼全长、体长及体重见表1。

表1 各组试验鱼的数量和规格
1.2 试验方法 试验参照GB/T18654.8-2008标准中耗氧率与临界窒息点的测定方法进行,在20℃ 温度下,分别测定花鮈骨、唇鮈骨及杂交鮈骨昼夜耗氧率变化。同时设计了15、20、25℃3个温度梯度,分别测定花鮈骨、唇鮈骨及杂交鮈骨的临界窒息点。
1.2.1 耗氧率的测定 耗氧率的测定采用流水式的密闭呼吸装置,参照GB/T18654.8-2008标准,同时对试验装置稍做改进,呼吸室容积为 10 L的广口瓶,呼吸室水温控制在20℃,水温变化幅度为±0.5℃。将试验用鱼放入呼吸室内,加盖并密封,排尽呼吸室内的空气,调节进水口流量,试验前需将试验鱼放在呼吸室适应2 h左右,待其呼吸平稳后开始试验,每隔2 h测定水流量和呼吸室进、出口水中的溶解氧,并记录水温,溶氧采用温克勒Winkler碘量法测定,取样3次进行测定,流速采用容量计时法测定,试验结束时用滤纸吸干鱼体表水分,用电子天平称量鱼体质量(精确到0.01 g),并测量鱼体全长(精确到0.1 cm)。
耗氧率计算公式如下:
式中:OC 为耗氧率,mg/g·h;D 为进水口的溶氧量,mg/L;DO为出水口的溶氧量,mg/L;V为试验过程中出水口平均水流速度,L/h;W为鱼体总重量,g。
1.2.2 临界窒息点的测定 测定装置同耗氧率测定装置,将试验鱼放入呼吸室,加盖密封并排尽呼吸室空气,试验前将试验鱼放呼吸室适应2 h左右,待其处于自然平静状态时,关闭进排水阀门,待呼吸室的50%试验鱼失去平衡、昏迷或濒临死亡时,测定水中的溶解氧,试验测定3次。
1.3 数据处理与分析 数据分析 试验数据利用Excel和 SPSS(19.0)软件进行统计分析,利用方差分析来检验温度对花鮈骨、唇鮈骨及杂交鮈骨耗氧率及窒息点的影响显著性。
2 结果与分析
2.1 耗氧率测定结果 在水温20℃条件下,花鮈骨、唇鮈骨及杂交F1代耗氧率的测定结果见表2。平均体质量(13.087±0.6959)g 的花鮈骨耗氧率平均值 为 0.1254 mg/(g·h), 平均体 质 量 (21.933±0.9952)g的唇鮈骨耗氧率平均值为 0.1099 mg/(g·h),平均体质量 (11.182±1.0452)g的杂交F1代耗氧率平均值为 0.1503 mg/(g·h)。
2.2 耗氧率的昼夜变化 由图1~图3可知,花鮈骨耗氧率昼夜变化较为平缓,在2:00时刻耗氧率最高,在 14:00时刻耗氧率最低,白天(8:00~18:00)耗氧率低于晚上(20:00 ~ 6:00);唇鮈骨耗氧率昼夜变化起伏较大,在2:00、4:00时刻耗氧率较高,耗氧率最低点出现在16:00时刻,白天(8:00 ~ 18:00)耗氧率低于晚上(20:00 ~ 6:00);杂交F1代耗氧率昼夜变化起伏不大,在2:00时刻耗氧率最高,在22:00时刻耗氧率最低,白天(8:00 ~ 18:00)耗氧率高于晚上(20:00 ~ 6:00)。
亲本及杂交F1代耗氧率的昼夜变化比较见图4所示,花鮈骨、唇鮈骨及杂交F1代耗氧率昼夜变化规律相似,其耗氧率最高点均出现在2:00时刻,亲本花鮈骨、唇鮈骨耗氧率最低点出现时刻较为接近,均出现在下午时刻,杂交F1代耗氧率最低点出现在晚间22:00时刻。唇鮈骨昼夜变化最为明显,其昼夜耗氧率差值为0.0357,昼夜耗氧率差值占平均值的32.48%。
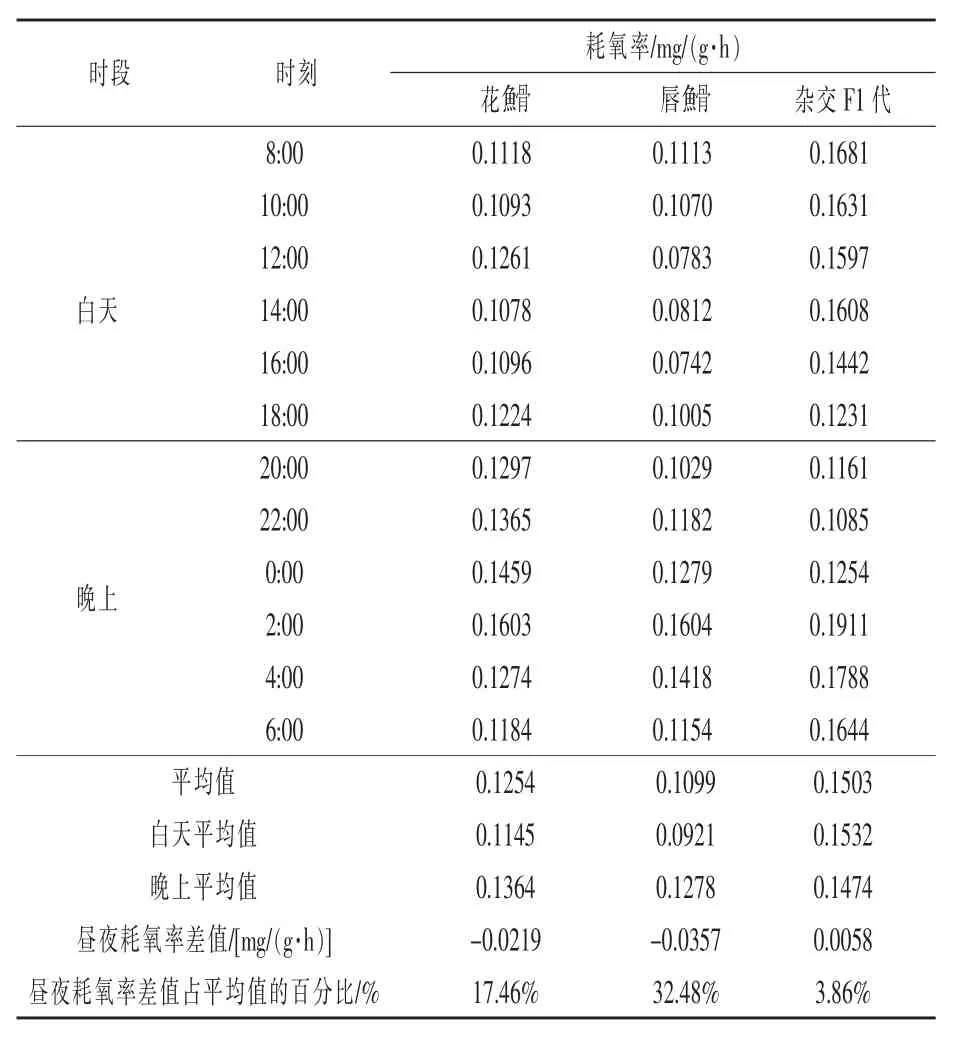
表 2 花鮈骨、唇鮈骨及杂交F1代耗氧率变化测定结果
2.3 临界窒息点 3种不同温度条件下亲本及杂交F1代窒息点测定结果见表3。到达临界窒息点时,花鮈骨、唇鮈骨及杂交F1代呼吸急促,部分鱼类向进水口集中,部分鱼类侧卧瓶底,鱼体逐渐翻白,腹部朝上,逐渐失去平衡,最后死亡。
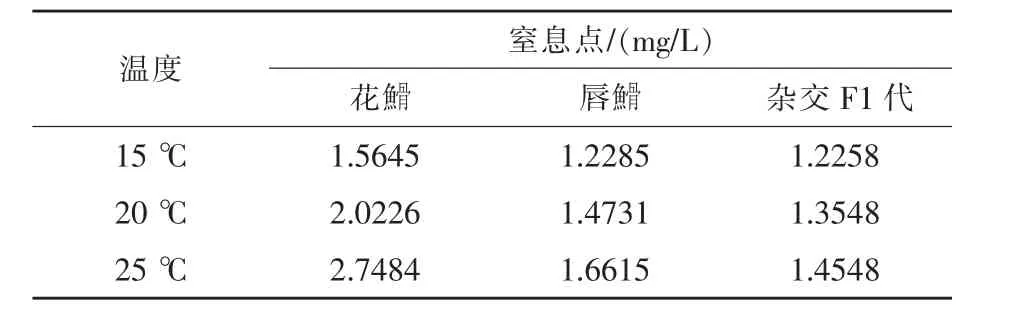
表3 花鮈骨、唇鮈骨及杂交 F1代窒息点变化测定结果
2.4 亲本及杂交F1代临界窒息点与温度的关系对表3数据分析花鮈骨临界窒息点与温度的关系,其临界窒息点随温度变化符合y=0.0054x2-0.0958x+1.7964的2次曲线关系 (R2=1)(如图5所示),在15~25℃时,其临界窒息点随温度上升而升高。
对表3数据分析唇鮈骨临界窒息点与温度的关系,其临界窒息点随温度变化符合y=-0.0011x2+0.0883x+0.1575的2次曲线关系 (R2=1)(如图6所示),在15~25℃时,其临界窒息点随温度上升而升高。
对表3数据分析杂交F1代临界窒息点与温度的关系,其临界窒息点随温度变化符合y=-0.0006x2+0.0461x+0.6648的2次曲线关系(R2=1)(如图7所示),在15~25℃温度范围内,其临界窒息点随温度上升而升高。
3 讨论
鱼类的耗氧率与种类有关,同时还与鱼类食性、生活习性、温度等有一定关系 (朱永久等,2014;杜劲松等,2011)。耗氧率高的鱼类预示着其活动相对活跃,可能在进行觅食或摄食等相关活动。鱼类的耗氧率变化规律有三种类型:白天耗氧高于夜间,此类鱼白天进食活动较为频繁;夜间耗氧高于白天,此类鱼属于昼伏夜出类型;白天与夜间耗氧相当,白天和夜晚都有进食。
本试验测定亲本及杂交F1代耗氧率的昼夜变化发现,花鮈骨、唇鮈骨及杂交F1代耗氧率昼夜变化均较明显,其中唇鮈骨耗氧率昼夜变化最为明显,其昼夜耗氧率差值为0.0357,昼夜耗氧率差值占平均值的32.48%;亲本花鮈骨、唇鮈骨白天耗氧率低于夜间,说明花鮈骨、唇鮈骨夜间活动也较为频繁,白天、夜间可能都有摄食活动;杂交F1代白天耗氧率显著高于夜间,属于白天进食频繁鱼类。生产实践中应根据鱼类代谢特性保持其适宜生长环境,保证鱼类正常生长。
花鮈骨、唇鮈骨及杂交F1代其临界窒息点随温度变化均符合2次曲线关系(R2=1),在15~25℃时,其临界窒息点随温度上升而升高。同一温度下,3种鱼类窒息点相比较,花鮈骨>唇鮈骨>杂交F1代,通过种类杂交选育,改善了父母本的生活习性,得到了在生长代谢方面具有优势的杂交子代,丰富了亲本及杂交F1代生长代谢、生理学理论,同时为F1代杂交优势研究、生产实践及应用推广提供了理论参考。
猜你喜欢
西南农业学报(2022年5期)2022-06-06
热带亚热带植物学报(2022年2期)2022-04-14
今日农业(2021年19期)2022-01-12
今日农业(2021年10期)2021-11-27
疯狂英语·初中天地(2021年6期)2021-08-06
广东农业科学(2021年3期)2021-04-23
儿童时代·幸福宝宝(2020年9期)2020-09-08
热带农业科技(2019年1期)2019-01-14
探索科学(2017年5期)2017-06-21
小说月刊(2014年11期)2014-04-18
