
补喂瘤胃保护性精氨酸对绵羊血浆中氨基酸浓度、激素水平和抗氧化能力的影响
2020-10-10 02:28:14王世昌张文杰侯传祥李晓斌张仕琦杨开伦
中国饲料 2020年17期
王世昌, 张文杰, 韩 明, 侯传祥, 李晓斌, 马 晨, 潘 军, 张仕琦, 杨开伦
(新疆农业大学动物科学学院,新疆肉乳用草食动物营养重点实验室,新疆乌鲁木齐830052)
瘤胃保护性精氨酸(RP-Arg)是物理保护的精氨酸,因其在瘤胃内具有良好的稳定性,引起了人们的关注(Chacher等,2012;李金霞等,2011),研究瘤胃保护性精氨酸在绵羊生产中的适宜添加水平,对于指导其科学利用具有重要意义。精氨酸对改善动物心血管功能、减少超氧化物产生以及促进激素分泌等方面具有良好作用 (Patel等,2017;Wu 等,2009),可用于反刍动物的营养调控剂。精氨酸极易被反刍动物瘤胃内的微生物降解,直接在日粮中添加,将会导致大量浪费(Chacher等,2012),限制了其在动物体内的利用率。目前,瘤胃保护性精氨酸在母畜上的研究取得了积极进展,主要集中在提高母畜的生长性能和繁殖性能方面。Yao(2008)给仔猪补喂精氨酸,结果表明,精氨酸显著增加了仔猪的平均日增重。饲粮中添加精氨酸可提高42~56日龄肉鸡的平均采食量和平均日增重(Corze 等,2003)。 赵宏丽(2013)给羯羊补喂瘤胃保护性精氨酸后发现,其可以改变肠道免疫性能和形态结构,维护肠道健康。给妊娠期母猪补喂精氨酸发现,窝产仔数、平均窝重、产活仔数分别提高17.91%、14.21%、22.87%(高开国等,2008)。但是,对补喂瘤胃保护性精氨酸后反刍动物机体中精氨酸代谢及其引起代谢变化的情况还需进一步研究。本试验旨在研究补喂不同水平瘤胃保护性精氨酸对绵羊血浆中氨基酸浓度、激素水平和抗氧化能力的影响,探讨瘤胃保护性精氨酸在绵羊生产中的饲喂效果,为其在绵羊生产中的科学应用提供参考依据。
1 材料与方法
1.1 试验设计 试验选取12月龄平均体重为(35.73±3.24)kg的新疆美利奴羊母羊 24只,随机分为4个处理组,每组6只羊,分别为对照组、试验Ⅰ组、试验Ⅱ组、试验Ⅲ组,试验期21 d。
试验羊基础日粮参照NRC(2007)绵羊的营养需要进行配制,并在日粮基础上,每组按照0、0.25、0.50、0.75 g/kg BW进行瘤胃保护性精氨酸的补喂。精料补充料组成及营养水平见表1,玉米青贮和苜蓿干草的营养水平见表2,精料补充料、玉米青贮、苜蓿干草中氨基酸含量见表3。
1.2 试验材料 瘤胃保护性精氨酸 (RP-Arg)购自辽宁亚禾营养科技有限责任公司,有效含量≥50%,过瘤胃率≥92%,水分≤10%。
1.3 试验时间与地点 本试验于2019年3月3日至2019年3月24日在新疆畜牧科学院中澳绵羊育种研究中心绵羊繁育试验基地进行。
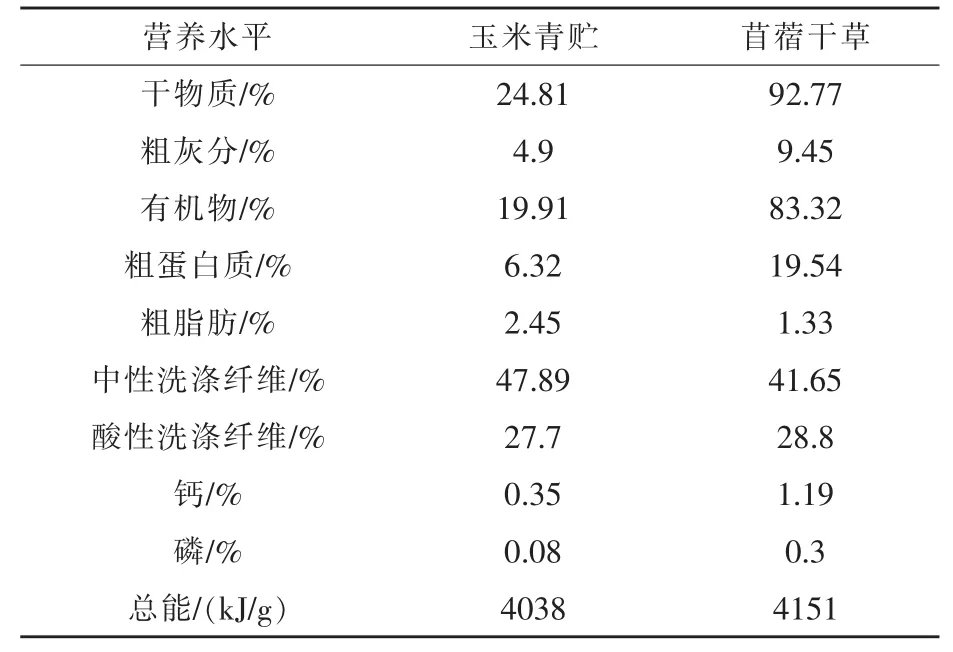
表2 TMR组成及营养水平(风干基础)
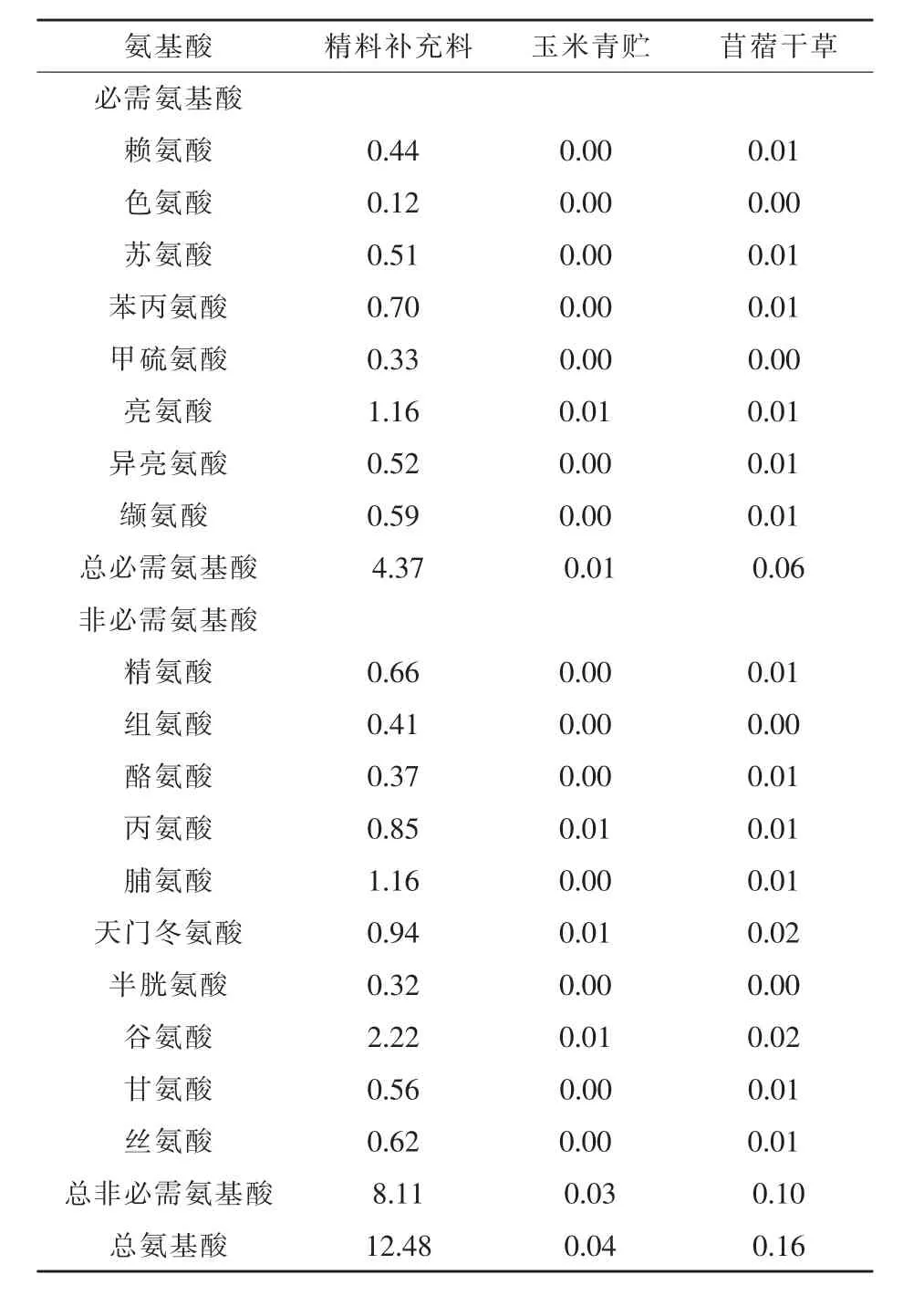
表3 精料补充料、玉米青贮、苜蓿干草中氨基酸含量(风干基础)%
1.4 饲养管理 全部试验羊在相同环境下进行统一饲养管理,自由饮水,将试验羊基础日粮和瘤胃保护性精氨酸平均分为两份,分别在每天08:00和20:00饲喂,其中瘤胃保护性精氨酸于每次饲喂前通过口腔灌喂的方式进行补喂,操作方法是将瘤胃保护性精氨酸放入装有50 mL普通饮用水的瓶中,人工口腔灌喂,灌喂完毕后用50 mL水润洗瓶壁,再次灌喂,确保瓶中无残留瘤胃保护性精氨酸。对照组采用与试验组相同的灌喂方式,仅灌喂等量的普通饮用水,以减小试验羊造成的误差。
1.5 样品的采集与处理 试验羊第21天晨饲前和晨饲后2 h进行颈静脉采血,并置于肝素钠抗凝管中,3500 r/min离心15 min,制备血浆,取上清液,分装至1.5 mL离心管,放入-20℃冰箱中保存。
1.6 样品的测定方法 试验饲粮中营养水平测定方法分别为粗蛋白质(CP)采用凯氏定氮法进行测定,中性洗涤纤维 (NDF)和酸性洗涤纤维(ADF)采用范氏洗涤纤维分析法测定,粗脂肪(EE)采用 GB/T6433-94 索氏抽提法测定,钙(Ca)采用邻甲酚酞比色法测定,磷(P)采用钒钼酸铵比色法测定,精料补充料、玉米青贮、苜蓿干草中氨基酸含量除色氨酸采用高效液相色谱法测定外,其余氨基酸采用GBT18246-2000方法测定。
绵羊晨饲前血浆样品送至北京华英生物技术研究所进行测定,血浆中尿素(Urea)、一氧化氮(NO)、丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、总抗氧化能力(T-AOC)、谷胱甘肽过氧化物酶(GSH-PX)测定使用比色法(A6半自动生化仪,北京松上技术有限公司),胰岛素(INS)、生长激素(GH)测定使用酶免法(华卫德朗DR-200BS酶标分析仪,无锡华卫德朗仪器有限公司)。
绵羊晨饲后2 h血浆样品送至天津普瑞华盛医学检验有限公司进行测定,血浆中氨基酸测定使用高效液相色谱-四级杆离子阱串联质谱仪(HPLC-LCMS/MS API3200 Q-TRAP, 美 国 AB SCIEX公司)。
1.7 数据分析 试验数据采用Excel 2010进行初步整理,使用SAS 8.1软件进行单因素方差分析,各组间采用Duncan’s法进行多重比较,试验数据用平均值表示,以P<0.05为差异显著水平,P<0.01为差异极显著水平。
2 结果
2.1 补喂不同水平瘤胃保护性精氨酸对绵羊血浆中氨基酸和尿素浓度的影响 由图1可知,将瘤胃保护性精氨酸补喂水平与绵羊血浆中精氨酸(A)和鸟氨酸(B)做回归分析,设瘤胃保护性精氨酸补喂水平为x,设绵羊血浆中精氨酸和鸟氨酸浓度为y,得到瘤胃保护性精氨酸补喂水平与绵羊血浆中精氨酸和鸟氨酸浓度的线性方程及非线性回归方程。绵羊血浆中精氨酸浓度随着瘤胃保护性精氨酸补喂水平的增加,呈线性升高的趋势(y=208.07x+149.27,R2=0.9863)。 绵羊血浆中鸟氨酸浓度随着瘤胃保护性精氨酸补喂水平的增加而升高,当补喂水平达到0.50 g/kg BW时,绵羊血浆中鸟氨酸浓度达到稳恒 (y=221.76+87.75 Inx,R2=0.8208)。结果显示,瘤胃保护性精氨酸补喂水平与绵羊血浆中精氨酸和鸟氨酸浓度之间具有较高的相关性,说明补喂瘤胃保护性精氨酸能提高精氨酸在动物机体内的利用率。
由表4可知,在必需氨基酸方面,与对照组相比,试验Ⅲ组绵羊血浆中赖氨酸浓度升高了24.02%(P < 0.05);试验Ⅰ、Ⅱ、Ⅲ组绵羊血浆中色氨酸浓度分别降低了 21.36%(P<0.05)、33.31%(P < 0.05)、27.02%(P < 0.05);试验Ⅱ、Ⅲ组绵羊血浆中苏氨酸浓度分别降低了26.23%(P< 0.05)、36.75%(P < 0.01);试验Ⅱ、Ⅲ组绵羊血浆中苯丙氨酸浓度分别降低了24.91%(P<0.05)、25.60%(P < 0.05);试验Ⅱ组绵羊血浆中甲硫氨酸浓度降低了31.19%(P<0.05);补喂不同水平的瘤胃保护性精氨酸对绵羊血浆中亮氨酸、异亮氨酸、缬氨酸没有显著影响(P>0.05)。
在非必需氨基酸方面,与对照组相比,试验Ⅰ、Ⅱ、Ⅲ组绵羊血浆中精氨酸浓度分别升高了36.33%(P > 0.05)、60.68% (P < 0.05)、107.09% (P <0.01),试验Ⅲ组比Ⅰ组升高了 51.91%(P<0.01);鸟氨酸浓度分别升高了 83.88%(P<0.05)、250.22%(P < 0.01)、264.15%(P < 0.01),试验Ⅱ组和Ⅲ组比Ⅰ组分别提高90.46%(P<0.01)、98.04%(P < 0.01);试验Ⅰ、Ⅱ、Ⅲ组绵羊血浆中酪氨酸浓度分别降低了17.63%(P<0.05)、24.70%(P < 0.01)、25.89%(P < 0.01);试验Ⅰ、Ⅲ组绵羊血浆中丙氨酸浓度分别降低了15.36%(P<0.05)、15.50%(P < 0.05);试验Ⅱ、Ⅲ组绵羊血浆中脯氨酸浓度分别降低了 24.68%(P<0.05)、20.85%(P<0.05);试验Ⅱ组比Ⅲ组绵羊血浆中天门冬氨酸高53.07%(P<0.05);补喂不同水平的瘤胃保护性精氨酸对绵羊血浆中瓜氨酸、组氨酸、半胱氨酸、谷氨酰胺、天门冬酰胺、谷氨酸、甘氨酸、丝氨酸没有显著影响(P>0.05)。

表4 补喂不同水平瘤胃保护性精氨酸对绵羊血浆中氨基酸和尿素浓度的影响
在尿素浓度方面,与对照组相比,试验Ⅲ组绵羊血浆中尿素浓度降低了22.93%(P<0.05)。
2.2 补喂不同水平瘤胃保护性精氨酸对绵羊血浆中激素水平的影响 由表5可知,与对照组相比,试验Ⅰ、Ⅱ、Ⅲ组绵羊血浆中胰岛素浓度分别升高了 13.59%(P > 0.05)、16.62%(P < 0.05)、28.74%(P<0.01),生长激素浓度分别升高了54.90%(P<0.01)、49.02%(P < 0.05)、41.67%(P < 0.05)。
2.3 补喂不同水平瘤胃保护性精氨酸对绵羊血浆抗氧化能力的影响 由表6可知,与对照组相比,试验Ⅰ、Ⅱ、Ⅲ组绵羊血浆中总抗氧化能力浓度分别升高了 31.76%(P < 0.01)、23.05%(P <0.01)、15.95%(P < 0.01);试验Ⅰ组绵羊血浆中超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶浓度分别升高了 24.64%(P < 0.01)、35.34%(P <0.01)、11.48%(P < 0.05), 丙二醛浓度降低了18.80%(P<0.05);试验Ⅱ组绵羊血浆中超氧化物歧化酶浓度升高了14.18%(P<0.05)。
与组间相比的结果表明,试验Ⅰ组和Ⅱ组绵羊血浆中总抗氧化能力、超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶、丙二醛、一氧化氮浓度均差异不显著(P>0.05);试验Ⅰ组比Ⅲ组绵羊血浆中总抗氧化能力、超氧化物歧化酶、过氧化氢酶分别高 13.64%(P < 0.01)、22.70%(P < 0.01)、25.94%(P<0.05),但绵羊血浆中谷胱甘肽过氧化物酶、丙二醛、一氧化氮浓度差异不显著(P>0.05);试验Ⅱ组比Ⅲ组绵羊血浆中超氧化物歧化酶浓度高12.40%(P<0.05),而两组间绵羊血浆中总抗氧化能力、过氧化氢酶、谷胱甘肽过氧化物酶、丙二醛、一氧化氮浓度差异均不显著(P>0.05)。
3 讨论
3.1 补喂不同水平瘤胃保护性精氨酸对绵羊血浆中氨基酸和尿素浓度的影响 在本试验中,绵羊补喂不同水平的瘤胃保护性精氨酸后,血浆中精氨酸和鸟氨酸显著升高,瓜氨酸浓度没有显著影响,这与前人研究结果一致。Davenport(1995)报道在羔羊日粮中补喂0.50、0.75 g/d BW瘤胃保护性精氨酸,结果显示,羔羊血浆中精氨酸、鸟氨酸浓度显著升高,瓜氨酸无显著变化;同样在母羊发情周期的第8~13天,母羊分别补喂0、90、180、360 mg/kg BW瘤胃保护性精氨酸,结果表明,补喂360 mg/kg BW瘤胃保护性精氨酸可显著增加血清中精氨酸浓度 (Saevre等,2011);Li(2014)研究表明,母猪妊娠第14~25天在饲粮中添加0.4%和0.8%的精氨酸可显著提高妊娠母猪子宫动脉血浆中精氨酸、鸟氨酸浓度,而对瓜氨酸浓度无显著影响;Agarwal(2017)在6周龄雄性大鼠基础日粮中分别添加2.5、7.5、12.5 g/kg的精氨酸,结果发现血浆中精氨酸浓度显著升高。精氨酸分解代谢涉及小肠、肝脏及肾脏等多个器官(Wu等,2007),其进入机体后大约40%在通过肠腔时被吸收,剩余约60%会被释放进入静脉血中(Wu等,1998),而高达70%的精氨酸在通过肠腔时转化为鸟氨酸(Kelley等,2014),鸟氨酸经鸟氨酸转氨酶在尿素循环中生成瓜氨酸 (Patel等,2017),瓜氨酸再经转化用于合成精氨酸,其中血浆精氨酸约5%~15%来自于血浆瓜氨酸的转化,剩余的血浆精氨酸来自于不同的来源,小肠中分解瓜氨酸的精氨酸琥珀酸合成酶(ASS)和精氨酸琥珀酸裂解酶(ASL)活性较低,因此,瓜氨酸在小肠中不能被分解,而是经小肠细胞释放到门静脉循环中,然后绕过肝细胞的代谢被运输到肾脏,最后在ASS和ASL催化下约80%的瓜氨酸被分解为精氨酸(Pons等,2017),这也解释了精氨酸、鸟氨酸浓度显著升高,瓜氨酸没有显著升高的原因。由此可见,外源添加瘤胃保护性精氨酸可以抵抗瘤胃微生物的降解作用,增加绵羊血浆中精氨酸和鸟氨酸的浓度。

表5 补喂不同水平瘤胃保护性精氨酸对绵羊血浆中激素水平的影响
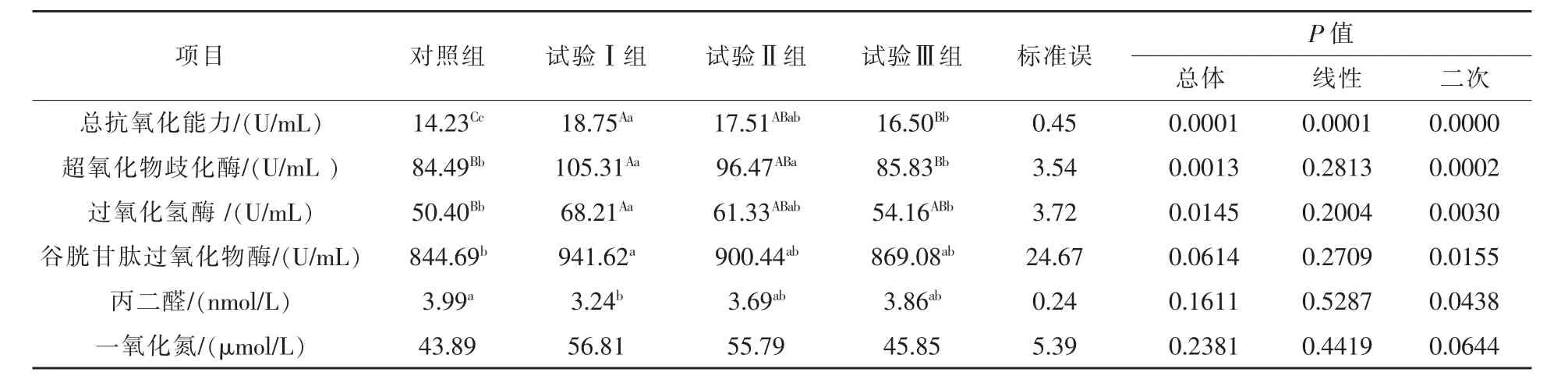
表6 补喂不同水平瘤胃保护性精氨酸对绵羊血浆抗氧化能力的影响
在必需氨基酸方面,绵羊补喂不同水平的瘤胃保护性精氨酸,血浆中赖氨浓度酸显著升高,色氨酸、苏氨酸、苯丙氨酸、甲硫氨酸浓度显著降低,亮氨酸、异亮氨酸、缬氨酸浓度没有显著变化。在非必需氨基酸方面,绵羊补喂不同水平的瘤胃保护性精氨酸,血浆中酪氨酸、丙氨酸、脯氨酸浓度显著降低,而对血浆中天门冬氨酸、瓜氨酸、组氨酸、半胱氨酸、谷氨酰胺、天门冬酰胺、谷氨酸、甘氨酸、丝氨酸无显著影响。这与Davenporta(1995)报道,在羔羊日粮中补喂瘤胃保护性精氨酸,结果显示除甲硫氨酸、缬氨酸外,其余必需氨基酸的研究结果一致;除丙氨酸、脯氨酸、组氨酸外,其余非必需氨基酸的研究结果一致。Li(2014)研究表明,母猪妊娠第14~25天在饲粮中添加0.4%和0.8%的精氨酸,子宫动脉血浆和尿囊液中亮氨酸、异亮氨酸、缬氨酸浓度的结果与本试验一致,而赖氨酸、色氨酸、苏氨酸、甲硫氨酸、苯丙氨酸、甲硫氨酸的结果与本试验相反;而非必需氨基酸精氨酸、鸟氨酸、瓜氨酸、酪氨酸、丙氨酸、组氨酸、半胱氨酸、天门冬酰胺、甘氨酸、丝氨酸与本试验结果一致,脯氨酸、天门冬氨酸、谷氨酸、谷氨酰胺结果不一致;马口服0.025%的精氨酸,颈静脉采血后结果显示,苏氨酸、苯丙氨酸、甲硫氨酸的结果与本试验一致,而赖氨酸、亮氨酸、异亮氨酸、缬氨酸的结果与本试验不一致,而非必需氨基酸精氨酸、鸟氨酸、瓜氨酸、丙氨酸、脯氨酸与本试验结果一致,组氨酸和谷氨酸结果与本试验不同(Kelley等,2014);Zeng (2008) 以雌性大鼠为研究对象,用丙氨酸作为等氮对照,在雌性大鼠日粮中补喂1.3%精氨酸盐酸盐后,血浆中亮氨酸、异亮氨酸、缬氨酸浓度的结果与本试验一致,赖氨酸、色氨酸、苏氨酸、苯丙氨酸、甲硫氨酸的结果与本试验不一致;而非必需氨基酸、精氨酸、鸟氨酸、瓜氨酸、组氨酸、谷氨酸、甘氨酸、丝氨酸浓度与本试验结果相同,血浆中酪氨酸、丙氨酸、脯氨酸、谷氨酰胺浓度与本试验结果不同。由此可见,在不同动物或试验条件下,补喂不同水平的瘤胃保护性精氨酸,动物绵羊血浆中氨基酸浓度的结果不同。
尿素主要由肝脏合成,可反映动物机体蛋白质分解代谢和肾功能情况(李忠荣等,2013)。在本试验中,补喂不同水平瘤胃保护性精氨酸使试验Ⅲ组绵羊血浆中尿素浓度显著降低,这分别与Zeng(2008)、Kim(2004)在仔猪、大鼠日粮中添加精氨酸的结果一致,可能是由于动物机体内氨基酸氧化的终产物是尿素,其浓度降低反映出机体对日粮中氨基酸利用率升高,当补喂瘤胃保护性精氨酸时,可减少机体内氨基酸的降解,最终减少血浆中尿素的浓度(杨平,2011)。但也有不一致的结果,Fligger(1997)给犊牛补喂 500 mg/kg BW 精氨酸血浆中尿素浓度显著增加。
3.2 补喂不同水平瘤胃保护性精氨酸对绵羊血浆中激素水平的影响 在本试验中,试验Ⅰ、Ⅱ、Ⅲ组补喂瘤胃保护性精氨酸可使血浆中胰岛素、生长激素的浓度显著升高,这分别与姚康(2008)、姜丽丽(2014)及麻名文(2009)在哺乳仔猪、雏鸭、肉兔日粮中添加精氨酸,血浆中胰岛素和生长激素浓度均显著升高的研究结果一致,与Zeitoun(2016)给妊娠早期和晚期的母羊分别补喂75、150 mg/kg BW的精氨酸,研究发现,补喂75 mg/kg BW精氨酸组的胰岛素水平高于150 mg/kg BW组和对照组,这与本试验结果一致。表明补喂瘤胃保护性精氨酸对血浆中胰岛素和生长激素具有促进作用。影响血浆中胰岛素和生长激素浓度变化的原因是胰岛β细胞在受到精氨酸的刺激后,带阳离子电荷的精氨酸通过在中性pH下通过质膜的直接去极化来实现胰岛素快速释放,生长激素浓度增加的同时也可刺激胰岛素的释放 (Philip等,2005;马晓静等,2003);当精氨酸抑制生长激素抑制素释放或脑垂体受到L-Arg刺激时,生长激素分泌增加(张迁,2018;Dasgupta 等,2006)。
3.3 补喂不同水平瘤胃保护性精氨酸对绵羊抗氧化能力的影响 在本试验中,补喂不同水平瘤胃保护性精氨酸,绵羊血浆中总抗氧化能力、超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶的浓度升高,丙二醛浓度降低,这与前人研究直接补喂精氨酸的结果一致。Liang(2018)和赵红霞(2018)研究表明,在大鼠及鱼日粮中添加不同水平的精氨酸,血清中总抗氧化能力、过氧化氢酶、超氧化物歧化酶、谷胱甘肽过氧化物酶浓度升高,丙二醛浓度降低。张浩(2016)选取妊娠期35 d,且怀有双羔的湖羊,饲喂50%NRC推荐量+20 g/d瘤胃保护性精氨酸,饲喂到妊娠110 d时,将妊娠羊屠宰采样,结果表明,妊娠羊胎盘中总抗氧化能力和超氧化物歧化酶浓度显著增加。总抗氧化能力是动物机体抗氧化系统状态的综合性指标,其浓度高低表明动物机体健康情况(张迁,2018)。超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶具有清除自由基的能力,丙二醛对细胞和哺乳动物的细胞具有致突变作用 (Marian等,2007)。超氧化物歧化酶可将有毒O2-分解为毒性较小的H2O2,H2O2是羟基(OH)自由基的潜在来源,过氧化氢酶和谷胱甘肽过氧化物酶将其转化为水,达到清除自由基的目的(Dasgupta等,2006)。可见,补喂瘤胃保护性精氨酸可以提高绵羊的抗氧化能力,增强机体抵御自由基的能力。
在本试验中,补喂不同水平的瘤胃保护性精氨酸对绵羊血浆一氧化氮浓度没有显著影响,这与 Zeng(2008)、杨慧(2012)、江雪梅(2011)在大鼠、妊娠母猪、母猪日粮中添加精氨酸,血浆中一氧化氮浓度显著升高的结果不一致。其原因可能是精氨酸的代谢产物之一胍丁胺,具有调节一氧化氮合成的作用(Piletz等,2013);精氨酸是合成一氧化氮的前体(Appleton,2002),而精氨酸与一氧化氮合成酶活性同时决定血浆中一氧化氮的浓度(郭洁平等,2013),本试验仅外源补充瘤胃保护性精氨酸,并不能增加血浆一氧化氮的浓度。
4 结论
补喂不同水平的瘤胃保护性精氨酸能显著提高绵羊血浆中精氨酸、鸟氨酸浓度,但使血浆中色氨酸、苏氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、丙氨酸、脯氨酸的浓度降低;能显著提高绵羊血浆生长激素、胰岛素的水平;还能显著提高绵羊血浆总抗氧化能力,当绵羊补喂剂量为0.25 g/kg BW时,效果最佳。
猜你喜欢
临床肝胆病杂志(2017年3期)2017-03-07 06:44:51
中国工程咨询(2017年7期)2017-01-31 03:01:26
中国工程咨询(2017年1期)2017-01-31 02:55:32
新农业(2016年15期)2016-08-16 03:40:48
西南军医(2016年3期)2016-01-23 02:17:56
中外医疗(2015年11期)2016-01-04 03:58:50
药学与临床研究(2015年4期)2015-06-05 11:35:53
中国当代医药(2015年30期)2015-03-01 02:08:13
现代农业(2015年5期)2015-02-28 18:40:57
湖泊科学(2014年2期)2014-02-27 10:30:05
