
D-半乳糖联合对氯苯丙氨酸致肾不藏志不寐大鼠下丘脑神经递质和凋亡基因的实验研究
2020-10-09 00:56任小娟张星平王庆全王冠英
中医药学报 2020年4期
任小娟,张星平,王庆全,王冠英
(新疆医科大学附属中医医院,新疆 乌鲁木齐 830000)
不寐病隶属于现代医学失眠症,其病因病机复杂,辨证思路多样,临床表现各异。中医治疗不寐病具有较好的临床疗效,但由于辨证论治的多样性导致其重复性较低。本课题组根据《黄帝内经》的五神理论、藏象理论以及后世医家相关理论,结合五脏辨证提出中医不寐五神分型诊断法[1]。该分型与五脏辨证相吻合[2],受中医师个体辨证倾向影响较小,客观性强,可操作性、可重复性强,也颇有疗效,便于临床、科研推广运用。具体包括肾不藏志不寐、心不藏神不寐、肺不藏魄不寐、肝不藏魂不寐和脾不藏意不寐。其中肾不藏志不寐为中医不寐五神分型中较为常见的证型,主症为夜寐早寤,病位在肾,其病因为阴、阳、气、血、痰、瘀、寒、热等各种原因导致肾脏的虚损,核心病机是肾志不入于舍,即肾不藏志[3]。
肾不藏志不寐是以早寤(早醒)为主要临床特征,多见老年人。这不仅与我们前期肾不藏志不寐临床研究相一致,而且与ANWAR E AHMED研究所发现的老年失眠多导睡眠图(PSG)中早醒的睡眠特征相吻合[4]。故我们选择衰老大鼠进行睡眠剥夺建立肾不藏志不寐大鼠。D-半乳糖致亚急性衰老和对氯苯丙氨酸(PCPA)致失眠是衰老研究和失眠研究中常用经典造模方法[5-6]。有研究发现抑制性神经递质γ-氨基丁酸(GABA)与睡眠有着密切联系[7];凋亡蛋白B细胞淋巴瘤基因-2(Bcl-2)和促凋亡因子Bcl-2相关X蛋白(Bax)不仅与衰老有关[8],还与睡眠密切相关[9]。因此,本实验采用D-半乳糖联合PCPA建立肾不藏志不寐大鼠,从神经递质和凋亡基因角度,观察该模型大鼠下丘脑神经递质和凋亡基因表达的变化,通过定位航行探索运动轨迹热图观察该模型大鼠的学习记忆能力,为肾不藏志不寐的进一步研究奠定实验基础。
1 材料与方法
1.1 实验动物及分组
SPF级雄性大鼠40只,体质量(200±20)g,由新疆医科大学动物实验中心提供,许可证号:SYXK(新)2018-0003。造模前将各组大鼠适应饲养1周后随机分成4组,每组10只,包括:空白对照组、老年组、失眠组和肾不藏志不寐组(模型组)。
1.2 主要药品与试剂
D-半乳糖,北京索来宝公司(批号:1013G051);对氯苯丙氨酸(PCPA),美国Sigma公司(批号:SHBJ7057);荧光定量PCR试剂盒由日本Takara公司提供。GABAARα1、GABAARβ2、GABAARγ2、Bax、Bcl-2引物由上海生工生物工程技术服务有限公司合成。引物序列见表1。

1.3 动物模型的建立
大鼠适应性饲养1周,然后每组大鼠予以对应处理,具体如下。
空白对照组:大鼠予以生理盐水120 mg/kg颈背部皮下注射,每日1次,连续42 d,然后生理盐水300 mg/kg腹腔注射,每日1次,连续3 d;老年组:大鼠予以D-半乳糖120 mg/kg颈背部皮下注射,每日1次,连续42 d,然后予生理盐水300 mg/kg腹腔注射,每日1次,连续3 d;失眠组:大鼠予以生理盐水120 mg/kg颈背部皮下注射,每日1次,连续42 d,然后PCPA 300 mg/kg腹腔注射,每日1次,连续3 d;肾不藏志不寐组:大鼠予以D-半乳糖120 mg/kg颈背部皮下注射,每日1次,连续42 d,然后PCPA 300 mg/kg腹腔注射,每日1次,连续3 d。
1.4 检测指标
1.4.1 大鼠体质量变化
造模结束后(第46天),测量各组大鼠体质量,观察各组大鼠体质量变化。
1.4.2 大鼠定位航行探索运动轨迹的比较
采用Morris水迷宫观察大鼠的空间学习记忆能力,利用水迷宫装置观察大鼠找到平台所需的时间,于第6天记录大鼠找到目标平台的定位航行探索运动轨迹,比较各组大鼠定位航行探索运动轨迹。
1.4.3 下丘脑GABAARα1、GABAARβ2、GABAARγ2mRNA相对表达量的测定
造模结束了,水合氯醛麻醉大鼠,冰上取大鼠脑组织,分离下丘脑,提取mRNA用于RT-PCR实验。RNA提取及cDNA合成参照试剂盒说明书进行,合成cDNA进行PCR扩增。采用 Primer 3软件设计 RT-PCR 引物,引物由上海生工生物工程技术服务有限公司合成。PCR 扩增条件:95 ℃预变性10 min,95 ℃变性10 s,退火10 s(退火温度:GABAARα1亚单位为60 ℃、GABAARβ2亚单位为60 ℃、GABAARγ2亚单位为60 ℃),72 ℃延伸10 s,40个循环后,95 ℃延伸15 s,60 ℃延伸1 min。将目的基因扩增结果经内参照校正后,以空白对照组目的基因扩增结果作为对照,2-△△ct法表达为mRNA的相对表达,计算其余各样本与空白对照组的比值,作相对定量分析,最终结果以相对定量结果表示。
1.4.4 下丘脑Bax、Bcl-2 mRNA相对表达量的测定
下丘脑cDNA合成步骤同上。采用 Primer 3软件设计RT-PCR引物,引物由上海生工生物工程技术服务有限公司合成。PCR 扩增条件:95 ℃预变性10 min,95 ℃变性10 s,退火10 s(退火温度:Bax亚单位为62 ℃、Bcl-2亚单位为62 ℃),余步骤同上。
1.5 统计学分析
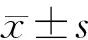
2 结果
2.1 体质量的比较
与空白对照组比较,模型组大鼠体质量明显减少(P<0.01;与模型组比较,失眠组、老年组大鼠体质量均明显增加(P<0.01)。见图1。
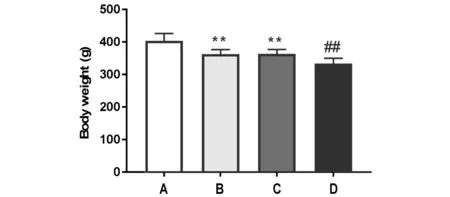
注:A:空白对照组;B:老年组;C:失眠组;D:模型组;与空白组比较,##P<0.01;与模型组比较,**P<0.01图1 各组大鼠体质量变化
2.2 定位航行探索运动轨迹的比较
与空白对照组比较,失眠组、老年组和模型组运动轨迹均明显延长,其中模型组找到目标靶平台运动轨迹最长。轨迹热图见图2。
2.3 下丘脑GABAARα1、GABAARβ2、GABAARγ2 mRNA的相对表达量
与空白组比较,模型组大鼠下丘脑GABAARα1、GABAARβ2、GABAARγ2mRNA的相对表达量均显著减少(P<0.05,P<0.05,P<0.01);与模型组比较,老年组大鼠下丘脑GABAARα1的相对表达量显著增加(P<0.05);与模型组比较,失眠组大鼠下丘脑GABAARβ2的相对表达量显著增加(P<0.05);与模型组比较,老年组和失眠组大鼠下丘脑GABAARγ2的相对表达量无明显变化(P>0.05)。见图3。
2.4 下丘脑Bax、Bcl-2 mRNA的相对表达量
与空白组比较,老年组、模型组大鼠下丘脑Bax mRNA的相对表达量显著增加(P<0.05),老年组、模型组大鼠下丘脑Bcl-2mRNA的相对表达量显著降低(P<0.05);与模型组比较,失眠组下丘脑Bax mRNA的相对表达量显著减少(P<0.05),失眠组大鼠下丘脑Bcl-2 mRNA的相对表达量显著增加(P<0.05);与模型组比较,老年组大鼠下丘脑Bax和Bcl-2 mRNA的相对表达量无显著变化(P>0.05)。见图4。
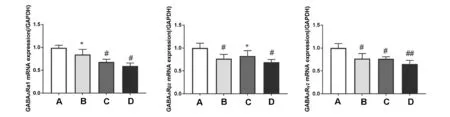
注:A:空白对照组;B:老年组;C:失眠组;D:模型组;与空白组比较,#P<0.05,##P<0.01;与模型组比较,*P<0.05图3 下丘脑GABAARα1、GABAARβ2、GABAARγ2mRNA的相对表达量
3 讨论
有研究发现老年人随着年龄的增加会伴随认知能力的下降[10],失眠会增加老年人痴呆的风险[11],这说明老年失眠会出现认知能力的下降。Morris水迷宫(MWM)实验可以分析动物的定位导航能力和空间探索能力,是推断实验动物的学习记忆能力的常用方法[12]。本实验发现肾不藏志不寐大鼠探索目标平台的运动轨迹明显长于正常大鼠,提示肾不藏志不寐大鼠学习记忆能力下降。
γ-氨基丁酸(GABA)是中枢神经系统中主要的抑制性神经递质,它能够抑制神经系统的神经元,具有神经保护作用,能够起到催眠、镇静和抗焦虑等功能[13]。GABA是通过与其受体相结合产生神经抑制性作用发挥镇静作用,其受体包括GABAA受体(GABAAR)、GABAB受体(GABABR)和GABAC受体(GABACR)[14]。其中GABAAR是离子通道型受体,是很多临床常用的镇静、抗焦虑及抗惊厥药等的靶受体[15]。GABAARα1、GABAARβ2、GABAARγ2型受体较常见,它们对睡眠的调节作用起着重要的影响[16]。JO K[17]研究发现黄精的水提取物能够增加大鼠皮质GABAARγ2蛋白表达从而改善睡眠。本研究发现肾不藏志不寐大鼠下丘脑GABAARα1、GABAARβ2、GABAARγ2mRNA的相对表达量均低于对照组,提示肾不藏志不寐的机制可能与下丘脑抑制性神经递质GABA基因表达下降有关。
睡眠具有保护神经元,防止和修复神经细胞损伤的作用[18]。B细胞淋巴瘤基因-2(Bcl-2)蛋白家族是第一个被发现参与凋亡调控过程的基因家族,在细胞凋亡过程中起着重要的调节作用[19]。Bcl-2家族成员由抗凋亡蛋白(Bcl-2,Bcl-2L和Bcl-w)和促凋亡蛋白(Bax,Bak,Bad,Bim和Bit等)组成[20]。其主要功能是直接调节线粒体膜的渗透性,调节促凋亡因子Bax的释放,从而发挥抗凋亡或促凋亡的作用。促凋亡成员和抑凋亡成员的比例可促进或抑制细胞凋亡的发生,其中以Bcl-2和Bax最具代表性[21]。有研究发现Bcl-2/Bax比例在晚上睡眠时间最高,而在失眠24 h之后发现Bcl-2/Bax比例会下降[22]。ARTAMOKHINAI V等发现Bcl-2、Bax在睡眠觉醒周期中也发挥着重要作用[23]。本研究发现肾不藏志不寐大鼠下丘脑Bcl-2 mRNA的相对表达量低于对照组,Bax mRNA的相对表达量高于对照组,提示肾不藏志不寐的机制可能与下丘脑神经细胞凋亡有关。
本实验研究了D-半乳糖联合PCPA建立的肾不藏志不寐大鼠下丘脑神经递质和凋亡基因。结果表明,肾不藏志不寐大鼠下丘脑神经递质基因GABAARα1、GABAARβ2、GABAARγ2mRNA的表达较正常对照组下调,下丘脑凋亡基因Bcl-2 mRNA的表达较正常对照组下调,Bax mRNA的表达较正常对照组上调。肾不藏志不寐大鼠模型的建立是肾不藏志不寐临床研究和实验研究的基础,神经递质和凋亡基因的调控可以作为肾不藏志不寐实验研究的新的切入点。
猜你喜欢
食品与生物技术学报(2022年1期)2023-01-11
中国图书评论(2022年4期)2022-11-22
世界科学技术-中医药现代化(2022年3期)2022-08-22
右江医学(2022年3期)2022-04-17
发酵科技通讯(2021年1期)2021-03-18
大众健康(2021年2期)2021-03-09
家庭百事通·健康一点通(2020年12期)2020-12-31
中老年健康(2017年9期)2017-12-13
校园英语·下旬(2016年4期)2016-05-09
安徽医科大学学报(2015年9期)2015-12-16
