
益生菌ABC转运体寡糖结合蛋白的结构学研究进展
2020-09-23 12:15苏思韵杨思婷张宇博周志立黄珍金钟先锋黄桂东
食品工业科技 2020年17期
苏思韵,杨思婷,张宇博,周志立,黄珍金,钟先锋,黄桂东
(佛山科学技术学院食品科学与工程学院,广东省传统发酵食品工程技术研究中心,广东省食品流通安全控制工程技术研究中心,佛山市酿造工程技术研究中心,佛山农业生物制造工程技术研究中心,广东佛山 528231)
人体肠道定殖一系列微生物菌群,共同维持着肠道的微生态平衡。益生菌是对人体健康有益的一类微生物,其包括双歧杆菌、乳杆菌、芽孢杆菌等[1]。这些菌群对宿主的益处表现在其能够产生维生素、氨基酸、短链脂肪酸等营养物质,有助于机体的代谢[2]。其次,益生菌能够产生抗菌肽等靶向作用于细菌的物质,从而有效地抑制病原菌[3]。同时,有助于增强机体的免疫应答[4],降低炎症反应[5-7]。益生菌还能参与治疗结肠炎[8]、自身免疫性疾病[9]、儿童咳嗽变异性哮喘[10]等疾病。
寡糖的摄入有助于益生菌的增殖和代谢,对人体有益。首先,寡糖的摄入有助于促进肠道益生菌的增殖和活力,显著增加双歧杆菌、乳酸杆菌、肠球菌的数量[11-13],同时有助于减少炎症及其相关症状[14]。其次,寡糖被益生菌代谢产生乳酸、短链脂肪酸等[15-17],降低肠道pH,抑制有害菌的生长。其能改善钙的吸收从而促进骨骼健康[18],还有助于降低心血管疾病风险[19-22]。
益生菌的基因组编码了一系列能够识别并转运寡糖的ABC通道[23-25]。其中一类ABC通道(如ECF型ABC通道),在摄入寡糖的过程中,不需要胞外底物识别亚基[24]。该通道由两个跨膜亚基(EcfS和EcfT)以及两个位于细胞质的亚基(EcfA1和EcfA2)组成。跨膜亚基EcfS能够直接识别底物,而EcfA1和EcfA2则是核苷酸结合亚基,能够组装成二聚体以协助供能。本课题组首先在嗜酸乳杆菌CICC22162中识别了转运水苏糖的ECF通道[24]。还有一类ABC通道(如MSM型ABC通道)[26],在摄入寡糖的过程中,需要胞外底物识别蛋白。该类型通道能够通过位于胞外区域的底物结合蛋白(solute-binding protein,SBP)识别寡糖底物,进一步通过SBP与跨膜亚基的互作将底物传递进细胞。
本文将益生菌结合寡糖的ABC通道胞外的底物结合蛋白称为寡糖结合蛋白。探讨寡糖结合蛋白识别寡糖的作用细节,有助于深入理解益生菌摄入寡糖的分子机制。经蛋白质结构学工作者的不懈努力,一系列ABC通道的寡糖结合蛋白的原子结构得以解析。本文将对益生菌中寡糖结合蛋白的结构特性及识别寡糖的异同进行综述。
1 ABC通道寡糖结合蛋白的共性
近年来,部分益生菌中寡糖结合蛋白的空间结构获得解析。与之前报道的ABC通道底物结合蛋白[27-29]相似,这些寡糖结合蛋白在结构上具有共性,具体表现在该类蛋白是由α和β两个结构域构成,在未与底物结合时,寡糖结合蛋白展示更加柔性的特征,使得α和β结构域能够在铰链区附近自由旋转。这种自由旋转使得蛋白的α和β结构域能够倾向于分开,从而使其构象以“开启”状态存在。在与底物结合时,蛋白构象以“关闭”状态存在,此时作为底物的寡糖被α和β结构域之间的接触面识别。该过程也被形象称为“捕蝇夹”机制[30]。图1A展示了典型的寡糖结合蛋白结构共性,包括了由铰链区连接的α和β结构域。图1B代表了结构已经鉴定的寡糖结合蛋白,分别是甘露寡糖结合蛋白(PDB:6I5W)[23]、阿拉伯木糖结合蛋白(PDB:4C1U)[31]、岩藻糖基乳糖结合蛋白(PDB:6HUR)[32]、乳糖-N-二糖结合蛋白(PDB:2Z8D)[33]、棉子糖系列寡糖结合蛋白(PDB:4RK9)。其α和β结构域之间形成底物的结合口袋,有利于底物的识别。
2 寡糖结合蛋白的结构特性及其底物功能研究
2.1 甘露寡糖结合蛋白
2.1.1 结构特性 双歧杆菌中存在能够结合甘露寡糖的蛋白BlMnBP1。该蛋白能够与甘露二糖、甘露三糖、甘露五糖发生相互作用。BlMnBP1与底物互作界面既包括直接的氢键相互作用,也包括水分子介导的间接氢键作用(如图2所示)。甘露二糖、甘露三糖和甘露五糖均存在于BlMnBP1的空腔口袋中,这三个寡糖具有不同的糖链长度。其中,甘露五糖在这三个寡糖中具有最长的糖链,能够与BlMnBP1的16个氨基酸形成氢键相互作用,而甘露三糖和甘露二糖则分别与BlMnBP1的10个和6个氨基酸形成氢键作用。甘露五糖能够与BlMnBP1的3个正电荷氨基酸K126、K152和K287形成最直接的盐桥相互作用,其盐桥的距离分别为5.35、5.46和3.68 Å。而甘露三糖和甘露二糖仅与BlMnBP1的1个正电荷氨基酸K287形成盐桥作用,其盐桥距离分别为3.75和3.80 Å。

图2 双歧杆菌甘露寡糖结合蛋白的空间结构信息
糖链最长的甘露五糖通过3个水分子介导与BlMnBP1的相互作用,而糖链最短的甘露二糖则通过13个水分子介导间接相互作用。其中Q223通过4个水分子与甘露二糖形成相互作用,而N163则通过2个水分子形成相互作用。这也表明短糖链的甘露二糖,需要通过更多的水分子,维持空腔处构象的稳定。
这三种寡糖与BlMnBP1之间形成微摩尔级别的结合力(表1)。SPR试验测得BlMnBP1与甘露二糖、甘露三糖和甘露五糖的结合力分别为187、82、108 μmol/L。ITC测得BlMnBP1与甘露二糖、甘露三糖的结合力分别为820和262 μmol/L。这表明甘露三糖在识别受体的过程中比甘露二糖的结合力稍强。甘露三糖识别BlMnBP1和BlMnBP2代表了一种相似的结合力。
2.1.2 底物的功能研究 甘露聚糖是一个重要的半纤维素聚合物,是木质纤维素的主要成分[34]。甘露聚糖能被肠道微生物,如青春双歧杆菌、嗜酸乳杆菌和盖氏乳杆菌等利用,而产气荚膜梭菌、大肠杆菌等产生有害物质的有害菌则不能利用甘露聚糖[35]。甘露聚糖被肠道菌群发酵后,最终生成乙酸、丙酸和正丁醇等短链脂肪酸[35],有利于调节肠道健康。
2.2 岩藻糖基乳糖结合蛋白
2.2.1 结构特性 双歧杆菌中存在能够结合岩藻糖基乳糖的蛋白BiFl2BP。BiFl2BP能够识别2′-岩藻糖基乳糖和3-岩藻糖基乳糖。与BlMnBP1和BlAXBP不同的是,岩藻糖基乳糖在与BiFl2BP相互作用过程中,除了氢键相互作用外,还通过范德华力与周围一系列非极性氨基酸形成疏水相互作用,如N39、A88和F92。这种疏水相互作用极大的增加了BiFl2BP与底物的结合能力。通过ITC及SPR对结合力的测定(表1)表明,BiF12BP能够以5~10 μmol/L的亲和力识别岩藻糖基乳糖,远高于BlMnBP1和BlAXBP对寡糖的识别。除了疏水相互作用,该蛋白能通过11至13个氢键相互作用识别底物,这极大的稳定了岩藻糖基乳糖靠近空腔的区域。特别是水分子介导的间接氢键相互作用中,2′-岩藻糖基乳糖能够通过水分子与Q63形成三个显著的相互作用,还能通过水分子与S212、S322形成间接相互作用。这些水分子介导的相互作用在3-岩藻糖基乳糖的识别过程中没有出现,也体现了该识别过程的特异性。
2.2.2 底物的功能研究 岩藻糖基乳糖是母乳寡糖的重要组成成分,能被长双歧杆菌摄入并产生乳酸和脂肪酸,显著降低肠道pH[36]。肠道中的病原菌如产气荚膜杆菌则不能大量利用岩藻糖基乳糖,且该菌的生长在低pH环境受到抑制[36]。体外实验表明,2′-岩藻糖基乳糖可抑制病原菌如空肠弯曲杆菌、肠致病性大肠杆菌、肠型血清沙门氏菌和铜绿假单胞菌对人肠细胞系Caco-2的粘附,而3-岩藻糖基乳糖可抑制人肠致病性大肠杆菌、铜绿假单胞菌对人肠细胞系Caco-2的粘附[37]。此外,2′-岩藻糖基乳糖能直接下调CD14的表达,影响革兰氏阴性菌的脂多糖的识别和结合,从而减少革兰氏阴性菌在肠道的定殖[38]。用含有2′-岩藻糖基乳糖的配方奶粉喂养婴儿,婴儿的炎症细胞因子较低,达到与母乳喂养的婴儿相似的水平[39]。
2.3 乳糖-N-二糖结合蛋白
2.3.1 结构特性 BlGL-BP对乳糖-N-二糖、半乳-N-二糖及乳糖-N-四糖的识别也展示了显著的差异。ITC测定BlGL-BP与这三个寡糖的相互作用(表1)表明,BlGL-BP能够较强地结合半乳-N-二糖,而与乳糖-N-四糖展示最弱的结合。结构分析表明(图3),BlGL-BP与这三个底物的结合表面出现了相似数量的氢键相互作用以及水介导的间接作用,特别是R49均能与三个底物形成盐桥。这些结构特性表明长双歧杆菌的乳糖-N-二糖结合蛋白的进化关系具有相近的同源性,这些蛋白对非还原性半乳糖的识别基本上是保守的,而相邻的位置是多样化,以适应不同的糖苷键和单糖残基[40]。

图3 乳糖-N-二糖结合蛋白与三个底物的作用细节
2.3.2 底物的功能研究 乳糖-N-二糖、半乳-N-二糖及乳糖-N-四糖是母乳寡糖的重要成分,在婴儿正常发育中发挥重要作用[41]。有研究[41-42]表明其能特异性地促进婴儿肠道的双歧杆菌增殖。
2.4 阿拉伯木糖结合蛋白
2.4.1 结构特性 双歧杆菌中还存在能够结合阿拉伯木糖的蛋白BlAXBP。BlAXBP与其底物的相互作用界面主要由氢键相互作用以及水分子介导的氢键作用组成(如图4所示)。在BlAXBP参与识别阿拉伯木三糖的过程中,共有10个氨基酸参与底物的直接氢键相互作用,分别是N39、S41、A42、N72、E73、F75、A76、L96、Q254和D386。其中,N39、S41、N72和G77也能够通过水分子介导与阿拉伯木三糖的间接相互作用。而在BlAXBP识别阿拉伯二糖的过程中,仅通过五个氨基酸N39、N72、Q254、Y346和D386与配体形成氢键相互作用。同时还可以观察到H199能够与配体形成直接的盐桥相互作用。水分子介导了8个间接的相互作用,这也表明二糖在结合空腔中的稳定存在也较多依赖于水分子。结合力测定(表1)表明阿拉伯木二糖和阿拉伯木三糖与BlAXBP的相互作用力较为接近,分别为81和71 μmol/L。
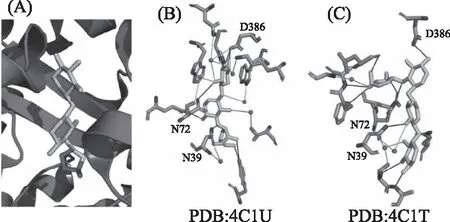
图4 双歧杆菌阿拉伯木糖结合蛋白的空间结构信息

表1 益生菌中已鉴定结构的寡糖结合蛋白
2.4.2 底物的功能研究 阿拉伯木聚糖(arabinoxylo-oligosaccharides,AXOS)具有促进人结肠双歧杆菌生长和生产丁酸盐的能力[43],具有双歧作用和产丁酸作用[44]。饮食健康人群每天摄入10 g AXOS,可以促进双歧杆菌的增长,同时抑制尿毒素对甲酚的排泄。另一研究表明,只需要在饮食中添加2.2 g的AXOS即可促进氮从尿道排泄转换到从粪便排泄,这种转换是一种有益的转换,减少了对肾的损伤[45]。长双歧杆菌(Bifidobacteriumlongum)和直肠真杆菌(Eubacteriumrectale)以AXOS为碳源共同培养时,两株菌株的生长和基因表达都受到共同培养的影响。长双歧杆菌消耗了AXOS的阿拉伯糖基,同时产生乙酸盐,乙酸盐随后通过直肠真杆菌的发酵产生丁酸盐[44]。
2.5 棉子糖系列寡糖结合蛋白
2.5.1 结构特性 芽孢杆菌中存在能够识别棉子糖系列寡糖家族的结合蛋白MsmE(图5)。其中,地衣芽孢杆菌中的BlMsmE与水苏糖的复合体结构模型在蛋白质数据库中已经报道。该结构模型代表了BlMsmE的一种“闭合”状态,即BlMsmE形成一个显著的结合口袋,与水苏糖形成直接作用。同时,枯草芽孢杆菌BsMsmE未与底物结合的空间结构也在数据库中得到报道,该模型采取了一种显著的“开启”状态。与“闭合”状态的BlMsmE不同,该开启状态的α结构域和β结构域倾向于分开,从而形成更加自由摆动的空间构象。这也意味着该类蛋白可能通过其显著的构象变化识别完成对棉子糖家族寡糖的摄入。

图5 棉子糖系列寡糖结合蛋白的晶体结构
2.5.2 底物的功能研究 棉子糖系列寡糖包括棉子糖、水苏糖、毛蕊草糖等。其中,水苏糖能改善肠道微环境[46],有助于糖尿病的治疗[47]。Liu等[47]比较了水苏糖和二甲双胍均对2型糖尿病大鼠的糖尿病有改善,同时降低大鼠的炎症水平。在分别给予大鼠水苏糖或二甲双胍4周后,采血分析生化指标。结果表明,水苏糖和二甲双胍均降低了血清脂多糖含量,同时降低炎症因子(IF-6)和肿瘤坏死因子α的表达。
2.6 环糊精结合蛋白
2.6.1 结构特性 干酪乳杆菌中存在环糊精和麦芽寡糖的结合蛋白(图6A)。该蛋白在与底物结合后,发生了显著的构象变化。该构象变化包括两部分,首先在螺旋处的位置发生了显著的空间迁移,其次部分螺旋区域变成了无规则卷曲。进一步比较LcMalE1与α-环糊精、β-环糊精、γ-环糊精的结合模式,发现α-环糊精形成了相对较小的环状构象(图6B),而β-环糊精和γ-环糊精展示了较大的环状构象,且这两个构象的尺寸较为接近。这很好的解释了LcMalE1与α-环糊精的结合力弱于β-环糊精与γ-环糊精。还比较了LcMalE1与β-环糊精和麦芽四糖的结合模式(图6C)。麦芽四糖在结合口袋的位置展示了与β-环糊精相似的非闭合环状模式,这种模式使其稳定的定位在结合口袋中,并且展示了与β-环糊精相似的强结合力。
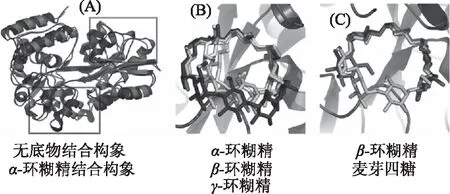
图6 乳杆菌寡糖结合蛋白的空间结构信息
2.6.2 底物的功能研究 环糊精在胃肠道中不能被消化,或只能部分消化,到达肠道后被肠道菌群发酵利用[48]。α-、β-、γ-环糊精分别由六、七、八个葡萄糖组成[49]。α-环糊精在肠道发酵产生有机酸、短链脂肪酸,通过调节脂类代谢相关基因的表达从而减少肥胖。Nihei等[50]研究了α-环糊精的摄入对高脂肪饮食诱发的肥胖模型鼠的影响。与高脂饮食小鼠肠道的拟杆菌、双歧杆菌和乳酸菌减少不同,摄入α-环糊精的小鼠肠道细菌总数增加。α-环糊精的摄入能够增加小鼠盲肠的乳酸和短链脂肪酸(如醋酸、丙酸和丁酸)的浓度,降低盲肠的葡萄糖浓度。此外,基因表达水平试验表明,α-环糊精诱导过氧化物酶体增殖物激活受体γ和α的基因表达上调,这两个受体分别与脂肪细胞的分化和能量消耗相关。而补充α-环糊精使固醇调节元件结合蛋白-1c和脂肪酸合酶的基因表达下调,该两个基因均与脂肪组织的脂肪酸和甘油三酸酯合成相关[50]。Grunberger等[51]研究表明,补充α-环糊精能改善肥胖II型糖尿病患者的脂肪代谢。患者连续3个月每天补充6 gα-环糊精,体重能减轻或至少保持在原来状态,对胰岛素敏感性增加,脂联素水平增加,血浆甘油三酯和低密度脂蛋白胆固醇降低[51]。
3 结论
胃肠道是一个复杂的生态系统,是一个多样化的高度进化的微生物群落[54]。肠道中的不同细菌有其偏好的摄食图谱,这种现象被称为“交叉摄食”[55-60]。有报道称,从婴儿肠道分离的长双歧杆菌偏好短链人体母乳寡糖,而不是复杂的高分子量人体母乳寡糖[61]。即使是同属于双歧杆菌的长双歧杆菌(B.bifidum)和婴儿双歧杆菌(B.infantis),其利用寡糖的策略都不同。B.bifidum和B.infantis存在于婴儿肠道中,竞争摄取利用母乳中的寡糖。B.infantis擅长通过寡糖结合蛋白摄入母乳寡糖,而B.bifidum使用几组糖基水解酶,先将复杂的寡糖降解,再摄入胞内[41]。与益生菌不同,肠道致病菌沙门氏菌的寡糖结合蛋白更偏向于结合单糖β-D-葡萄糖、β-D-半乳糖等单糖[62-65]。研究肠道不同菌群偏好的寡糖,有助于精准调控肠道菌群,促进人体健康。
ABC通道是益生菌重要的寡糖摄取通道,其寡糖结合蛋白的作用是识别和结合寡糖,很大程度上决定了ABC通道的底物特异性[31,66]。寡糖运输蛋白是优选的研究糖代谢偏好性的探针,因为它能特异性直接与特定寡糖作用[40]。寡糖转运体的生化和结构表征,是实现基于基因组序列数据的代谢偏好研究的关键[40]。此外,了解寡糖结合蛋白与底物的作用细节,有助于开发新型益生元,评价益生元的功效。随着经济社会的发展,功能性食品行业发展迅猛。当代功能性食品的开发,需要经过人体及动物实验验证是否具有某种生理功能。同时,还需要明确功效成分,及其含量、结构、作用机理等。研究益生菌的寡糖结合蛋白,以及该类蛋白的寡糖识别机制,为益生元类功能性食品的开发提供理论支撑。
本论文综述了人体肠道益生菌中ABC通道寡糖结合蛋白的结构学研究进展。尽管不同的寡糖结合蛋白展示了三级结构相似性,其特异性识别寡糖的分子机制各不相同。X-射线晶体结构揭示了寡糖结合蛋白识别线型、环型寡糖的原子细节,也揭示了其结合底物,以及未结合底物的结构异同。目前,还存在一系列益生菌的寡糖结合蛋白尚未被鉴定和解析,对其结构学研究为进一步认知益生菌寡糖摄入机制,提供研究基础。
猜你喜欢
食品与生物技术学报(2022年1期)2023-01-11
中国免疫学杂志(2019年17期)2019-09-20
现代食品(2018年22期)2019-01-28
中成药(2018年8期)2018-08-29
中国化肥信息(2018年6期)2018-08-23
中成药(2018年6期)2018-07-11
中成药(2018年4期)2018-04-26
中成药(2017年8期)2017-11-22
中成药(2017年5期)2017-06-13
中国当代医药(2015年31期)2015-03-01
