
脱水过程中蚕豆种子内部水分相态变化及表面水分损失动力学研究
2020-09-23 12:15:40李言洁王雅博王金山
食品工业科技 2020年17期
诸 凯,李言洁,王雅博,王金山
(天津商业大学,天津市制冷技术重点实验室,天津 300134)
蚕豆种子作为一种生命材料,与土豆、苹果等食品的干燥不一样,有种子发芽的要求,在保存时对种子活性要求高,收获后的蚕豆种子需要及时脱水到安全含水率才能长期储藏。并且因其脱水(传热和传质)在细胞层面所展现出的规律特征,对研究其他种子的长期存活也有一定借鉴作用。热风干燥设备简单,操作方便,但传热效率较低,产品内外干燥不均,易出现表皮硬化现象,不利于内部水分排出;微波干燥是从物料的内部开始加热,具有时间短、速度快的特点,但由于微波较强的穿透性,使物料内部水分快速迁移至表面,致使表面积存大量水分,造成样品内部较硬,表面焦化现象严重[1]。低温真空干燥是指物料在具有一定真空度的密闭容器内低温加热,使物料内部的水通过压力差或浓度差扩散到表面,而后被真空泵抽走的干燥方法[2]。既能避免食品物料在高温下引起的热力损伤,又能防止冰晶在冻结过程中产生的机械损伤,使物料呈现良好的结构性能。因此对于种子储存,多采用低温真空干燥方式将种子脱水到指定含水率进行保存。
低场核磁共振(low-filed nuclear magnetic resonance,LF-NMR)是利用氢原子核在磁场中的自旋弛豫特性[3],通过弛豫时间的变化从微观的角度解释样品中水分的分布变化和迁移情况,具有快速、准确、无损、无侵入等优点[4]。生命组织的细胞结构复杂,干燥过程中的内部结构和水分形态变化难以了解[5],为更直观地观察组织内部的水分流动特性,研究人员利用核磁共振技术,观察到植物组织内至少存在两种或以上的水分相态[6-11]。Almeida等[6]对硬木材分离出了三种不同形式的水分:导管分子中的液态水,纤维和薄壁组织中的液态水以及结合或细胞壁水;张绪坤等[7]将胡萝卜在热风干燥过程中的水分相态划分为自由水、半结合水和结合水;宋平等[8]发现水稻浸种过程中种子内部水分存在结合水、自由水2种水分状态,同时可区分出内层水、中层水、外层水3种水分分层;Xu等[9]检测到西兰花存在三种不同束缚大小的水分,分别是液泡和木质部中流动性较高的水分,细胞质中与大分子(如蛋白质)相互作用的流动性较低的水分以及与细胞壁多糖结合的束缚水;Khan等[10]对植物组织细胞间水、细胞内水和细胞壁水三种水环境下的水分进行了相态划分,胞间水称为毛细水或游离水,其束缚程度较弱的胞内水为松散束缚水(LBW),而细胞壁水则为强束缚水(SBW);然而在研究吸湿性食品在干燥过程中细胞膜破裂现象时,Khan等[11]又认为在不同的细胞环境中含有游离(FW)和结合(BW)水。目前,此技术已成功应用于肉质食品加工、谷物干燥、果蔬储存等的干燥过程中的水分相态及其变化形式,但对蚕豆种子真空脱水过程中水分存在相态尚无报道。
脱水是一种常用的果蔬、种子保存方法。然而,因为植物内部结构以及热质传递的复杂性,了解实际传热传质过程对优化能源效率和保持食材质量至关重要[12]。干燥过程中的水分和温度分布及其传输机制是非常重要的,而水分输送速率取决于细胞的大小和方向,以及样品中细胞水的类型[13]。因此本文将利用核磁共振技术观察蚕豆脱水过程中细胞层面的水分扩散行为,并数值模拟其传热传质过程,减少实验次数,降低成本,从而将微观层面的水分相态转变与宏观含水率变化相结合,研究蚕豆种子在低温真空环境下的温度分布、水分分布以及水分状态随时间的变化规律,以及环境条件对脱水的影响,为蚕豆种子真空干燥工业化生产和产品保藏研究提供理论依据。
1 材料与方法
1.1 材料与仪器
蚕豆 新鲜蚕豆样品在当地市场购买,在使用前在2~4 ℃条件下储存,初始湿基含水率为65%~70%,单粒约重2.352 g,长25.3 mm,宽17.4 mm,厚8.3 mm。所选样品大小均匀,无病。
NMI20-025V-I核磁共振成像分析仪 上海纽迈电子科技公司;FA2204C电子天平 分度值0.1 mg,上海越平科学仪器公司;T型热电偶 0.13 mm,日本;ST700压力变送器 美国霍尼韦尔;GM10数据采集器 日本恒河公司;XODC-1030A低温恒温器 南京先欧仪器制造有限公司;JK-100.4扩散真空泵(真空室由不锈钢筒体和平板组成,底部厚度设计为1 mm) 上海五洲真空设备有限公司。
1.2 实验方法
1.2.1 低温真空脱水实验 将被干燥的物料在高于物料共晶点温度下的低温环境中进行冻结,然后将其置于高真空环境中,使物料中的水分蒸发,从而将物料中的水分除去[14]。大多数蚕豆在-1 ℃左右的温度下开始结冰,当温度继续降低至约-5 ℃时,其70%~80%的水分结冰,蚕豆即随之变硬。在-5~-1 ℃的温度下,冰结晶大量的形成,称为蚕豆冰结晶最大形成阶段。若温度进一步降低至-35~-30 ℃,冻结过程随即完成。本实验选择0 ℃以上的几个温度作为蚕豆脱水温度,可以避免蚕豆内部水分结冰使内部组织结构受损,以及便于观察不同相态水分间的转化。将挑选好的蚕豆放入低温真空干燥腔中,通过低温恒温水浴装置设置腔内温度为0、4、8 ℃,调节真空泵控制阀使腔内真空度稳定在95、97、99 kPa。每脱水1 h后,用电子天平测量不同时刻(0~30 h)的蚕豆重量,计算此刻蚕豆种子的含水率。
1.2.2 核磁共振分析 设置核磁共振成像分析仪的检测温度,使其稳定在32 ℃。将脱水实验得到的蚕豆样品,立即放入放样管中进行检测。利用核磁共振波谱分析软件中的FID(free induction decay)脉冲序列寻找磁场的中心频率及硬脉冲脉宽,之后选取硬脉冲回波序列CPMG(carr-purcell-meiboom-gillsequence)测定蚕豆样品的横向弛豫时间T2[15],通过弛豫时间的变化从微观的角度解释蚕豆样品中的水分状态、含量变化和迁移方向。
1.3 数值模拟方法
1.3.1 几何建模及网格划分 根据图1(a)蚕豆种子实际形状,采用Pro ENGINEER 5.0建模软件对蚕豆种子三维重构,形成三维实体,从而建立蚕豆的几何模型,如图1(b)。图1(b)中五点为蚕豆不同位置点处坐标,长、宽、高方向分别用y、x、z轴表示,蚕豆中心点坐标为(0,0,0)。模型中x方向的最大宽度为19 mm,最小宽度为17 mm;y方向的最大长度为26.5 mm,最小长度为9 mm;z方向最大厚度在两头为10 mm,中间部位最薄为7.8 mm。将此三维几何模型导入多物理场耦合计算模拟软件COMSOL Multiphysics 5.4中进行网格划分,共包含764036个自由四面体网格,如图1(c)。
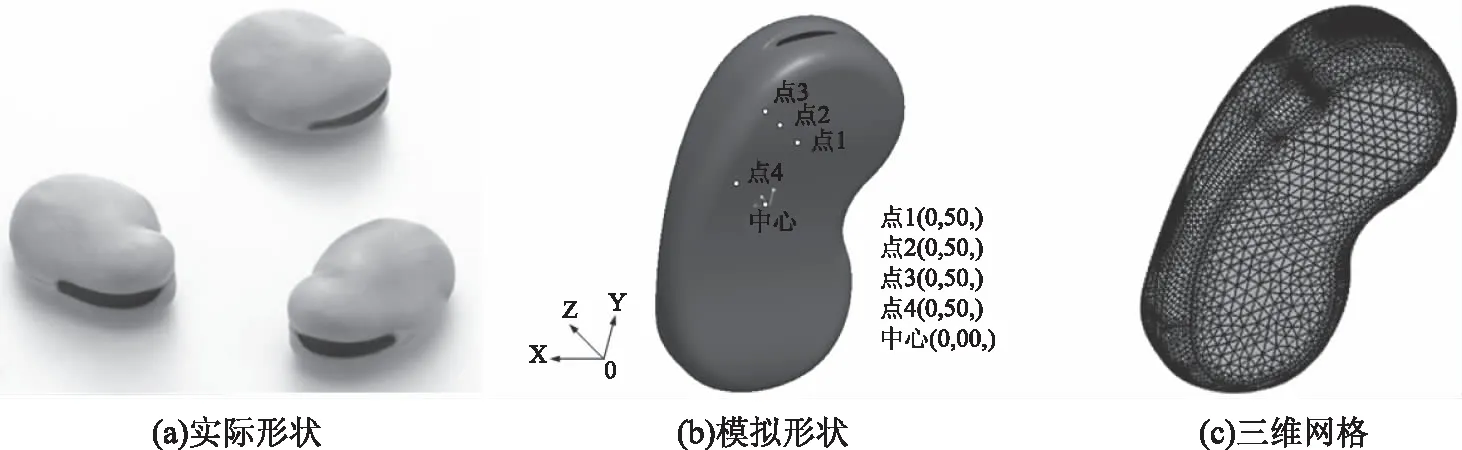
图1 几何模型建立
1.3.2 热湿传递模型及定解条件 为研究蚕豆在低温真空脱水过程中的热质传递机理以及其内部水分分布规律,根据生物多孔介质中温度、水分之间复杂的耦合关系,基于菲克扩散定律、傅立叶导热定律等,运用有限元法COMSOL Multiphysics 软件平台数值模拟蚕豆水分迁移过程。其中需要考虑以下两种物理效应:稀物质传递(tds)以及固体传热(ht)分别描述水分浓度和温度的瞬态物理场接口。此模型不对蚕豆外部的对流速度场进行建模,因为已给定与周围空气发生对流传热和水分传递的系数。模型基于以下假设:忽略变形不影响热质传递,因此当仅研究脱水过程中热质传递机理可以忽略蚕豆种子体积和外形的变化;初始时刻蚕豆内部水分和温度均匀分布;蚕豆物性为各向同性。
1.3.2.1 传热模型 控制方程:
式(1)
式中,ρeff-蚕豆有效密度,取平均值1210 kg/m3;cp,eff-蚕豆恒压比热容,取1859 J/(kg·K);T-蚕豆内部温度,K;t-时间,s;λeff-蚕豆的导热系数,实测取平均0.52W(m·K);Q-热源,W。
蒸发需要能量,由边界热源条件表征:
Q=-Hvaphd(cb-c)
式(2)
式中,Hvap-水的蒸发潜热,取2459.1 kJ/mol;hd-蚕豆的传质系数,采用实验数据拟合,范围0.0166~7.67×10-6m/s;cb-空气水分浓度,取1377.8 mol/m3;c-蚕豆任意时刻的水分浓度,mol/m3(水分浓度均以蚕豆的水分浓度为基础转换)。
边界条件:
式(3)
初始条件:
T|t=0=T0
式(4)
式中,h-蚕豆表面对流换热系数,取10 W/(m2·K)[16];Tb-冷空气温度,取0、4、8 ℃;T0-蚕豆初始温度,取25 ℃。
1.3.2.2 传质模型 控制方程:
式(5)
边界条件:
式(6)
初始条件:
c|t=0=c0
式(7)
式中,Deff-蚕豆的水分有效扩散系数,m2/s;c0-蚕豆初始水分浓度,取46844 mol/m3。
实验含水率数据采用以下公式进行无量纲化:
式(8)
式中,MR-无量纲含水率;Mt、Me和M0分别为任意时刻的干基含水率、脱水平衡时所对应的干基含水率以及初始时刻的干基含水率,kg水/kg干物质,可简化为Mt/M0。
有效水分扩散系数是衡量水分输送能力的重要参数。水分扩散是物料从中心到表面的主要传质机制[17]。基于菲克扩散第二定律,采用Crank(1975)的含水率与脱水时间的关系:
式(9)
式中,M为蚕豆平均含水率,F0表示传质过程中的傅立叶数,n为傅立叶数的项。此公式非常适合真空脱水,且内部传质阻力是控制整个传质过程的主要机制。水分有效扩散系数则可以表示如下[18]:
式(10)
式中,Deff为水分的有效扩散率;L表示特征长度(假设蚕豆为平板,厚度方向);t是脱水时间。通过实验可以得到L以及不同时刻的M,利用二分法由式(9)可以得到F0,其中n=60。然后再对F0求导,确定有效水分扩散系数随时间变化的规律。
2 结果与分析
2.1 核磁共振实验
2.1.1 NMR总信号幅值与纯水含量关系分析 确定纯含水量与核磁共振总信号幅值(峰面积总和)的关系,如图2所示。可以看出蚕豆水含量与NMR信号幅值呈显著的线性关系,并且可由回归方程求得脱水过程中各状态水分的含量。因此,通过分析核磁共振弛豫谱可以间接反映出蚕豆种子脱水过程水分的分布及迁移规律[19]。由于含水量与信号幅值之间呈线性关系,蚕豆水含量变化则可由核磁共振信号幅值定量表示,以便观察和分析。

图2 NMR总信号幅值与纯水含量拟合曲线
核磁共振横向弛豫谱(transverse relaxation spectrum),即T2弛豫谱,给出了T2弛豫谱信号幅值以及对应的 T2弛豫时间,其中前者与纯水含量有关,后者反映了水分相态信息[20]。不同化学微环境中的质子具有不同的横向时间[21],T2弛豫时间越短,质子的自由度越小,对应状态的水分越不易被排出;T2弛豫时间越长,质子的自由度越大,对应状态的水分越易被排出[22]。植物性食品材料具有多孔性和吸湿性,组织内包含细胞间水、细胞内水和细胞壁水三种水环境[5]。借鉴Sun等[23]对微波真空干燥过程中果蔬的水分状态划分方法,新鲜蚕豆一般有3~4个峰,较短弛豫时间范围T21(0.01~1 ms)对应与细胞内大分子结合的结合水,较长弛豫时间范围T22(1~50 ms)对应亲附于组织胶体表面的半结合水,以及最长弛豫时间范围T23(50~300 ms)对应游离在细胞间,自由度较高的自由水。
蚕豆在温度为4 ℃、真空度为97 kPa的环境中脱水,并利用核磁共振技术检测其水分含量与状态。将脱水不同时长的T2弛豫谱放到同一坐标轴上,可以观察到其水分变化的过程,如图3所示。显然,随着脱水时间的延长,信号幅值总是往低横向弛豫时间方向迁移,最终趋于零。这是由于自由度高的水分不断蒸发并减少,使整个蚕豆的水分状态逐渐趋于自由度低的状态。

图3 蚕豆在4 ℃、97 kPa条件下T2弛豫谱随脱水时间的变化
2.1.2 蚕豆内部水分状态变化规律 图4为三种水分状态占比随脱水时间变化,图5是各状态水分对应的信号幅值随脱水时间变化。从图4、图5中可以看出在低温真空脱水过程中,总体水含量不断减少,各相态水分含量也在不断变化。自由水含量最多,因其自由度高,在脱水开始后迅速向外排出,一直不断减少,约12 h蒸发完。半结合水在自由水大部分被脱除前不会向外逸出,且在前8 h还会缓慢增多,这是由于葡萄糖、果糖等碳水化合物浓度的升高,小部分自由水向半结合水转化,另一方面,随着脱水的进行,物料内营养物质以及酶分解出的结合水也会转化为半结合水[24]。同时,由于自由水的减少,也使得半结合水占比在前12 h内持续上升,在此之后开始减少,在t=26 h时含量减少了88.5%。结合水占比虽然一直在增大,但受到的束缚最强,为维持细胞内外水分浓度平衡,含量仅小幅度波动变化,到t=26 h时含量减少了45.3%。

图4 各水分状态含水比率随脱水时间变化

图5 三种水分状态水含量随脱水时间变化
2.2 数值模拟
2.2.1 有效水分扩散系数的数值估计 蚕豆水分扩散系数Deff随时间变化的结果如图6所示。与预期一样,真空和温度对Deff的影响是显而易见的。Deff值先迅速增大,达到最大值,然后随时间逐渐减小。在高真空环境下,随着温度的升高,Deff的变化更加明显。这是因为当真空度较高时,水分更容易变成水蒸气,导致Deff更易增大,而当脱水时间超过8 h后,两个因素的影响都较弱。实验得到的平均有效水分扩散系数为0.65~2.97×10-10m2·s-1,与Zogzas等[25]分析的生物材料有效水分扩散系数范围在10-9~10-11m2·s-1内的结果一致。然后将计算得到的水分扩散系数结果作为参数输入模型中,对蚕豆脱水过程进行模拟。

图6 蚕豆脱水过程中有效水分扩散系数随时间的变化
2.2.2 模型验证 模型验证主要通过比较试验和模拟蚕豆脱水曲线。图7表示温度为4 ℃、真空度为97 kPa时,初始温度为25 ℃的蚕豆脱水实验和数值模拟的对比曲线。从图7中可看出,蚕豆实验曲线和模拟脱水曲线趋势一致,两者之间最大相对误差为15%,低于一般数值模拟的20%精度要求,因此数学模型得到验证。

图7 模拟与实验蚕豆脱水曲线
2.2.3 蚕豆在低温真空环境的温度分布 模拟了初始温度为25 ℃(298.15 K)的蚕豆,在温度为4 ℃、真空度为97 kPa的环境下的脱水过程,其脱水初期的温度分布及各内部点(位置示意图见图1(b))处温度变化曲线,如图8、图9所示。在脱水初期,蚕豆表面与外部环境存在压力差,导致水分扩散系数急剧增大,水分迅速蒸发,带走大部分热量,使得蚕豆温度急剧下降。因此在0.7 h内蚕豆温度下降了约25 K,之后水分扩散减慢,蚕豆与空气之间对流换热,温度缓慢上升3~6 K,直至达到冷空气温度277.15 K。从图9中可以看出,外部点4温度变化最大可达26.7 K,内部点温度变化最小,这表示蚕豆边界点较中心位置降温快,导致在脱水初期内部会存在温度梯度。在t=0.3 h时刻内部点和外表面温差可达17 K,随着热量的传递,温差逐渐减小,直至为零,使整个蚕豆温度均匀且与外部空气温度一致。

图8 不同时刻蚕豆内部温度分布
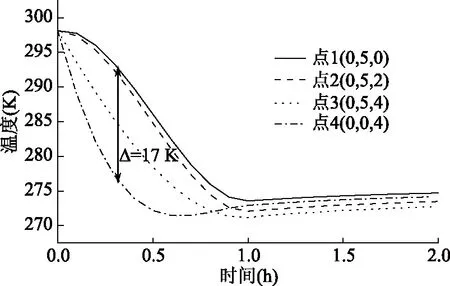
图9 蚕豆各内部点温度变化曲线
2.2.4 蚕豆在低温真空环境的水分分布 图10为蚕豆在温度为4 ℃、真空度为97 kPa的模拟环境下,不同时刻(t=5,10,15,20,25 h)中心点处XY截面的蚕豆内部水分分布图。可以看到,蚕豆内部水分在两头各为中心向外扩散。这是因为蚕豆中间部分比较薄,而上下两头比中间凹陷部分更厚,当蚕豆表面的水分向周围空气逸出后,由于厚度不均,其内部水分浓度也呈不均匀分布。蚕豆种子内部的水分在初始时刻呈均匀分布,随着脱水的进行,水分通过边界不断向周围环境蒸发,使得整个种子呈现中心含水量相对更高的水分不均匀分布状态。
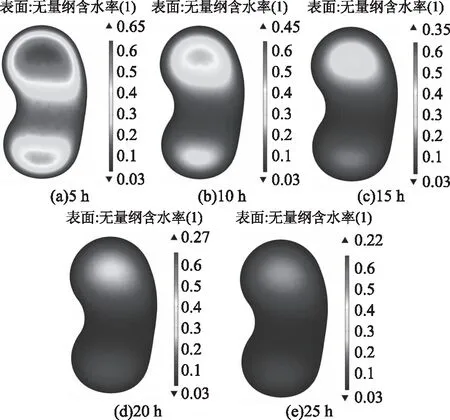
图10 不同时刻蚕豆内部水分分布
图11是蚕豆内部点含水率模拟变化曲线。显然,外部点4比内部点1脱水更快,在t=3 h时含水率相差约0.65。与核磁共振水分相态结合分析可知,在脱水过程的前10 h内,自由水迅速蒸发;在10~15 h期间,自由水蒸发完全,大量半结合水开始蒸发,但蒸发速度较慢,这是由于整体含水率较低,内外浓度差较小,导致脱水缓慢;而在15~30 h内,结合水和剩余的小部分半结合水极其缓慢地蒸发,蚕豆含水率仅略有降低。在真空脱水过程中,物料表面水分蒸汽与干燥腔内蒸汽存在压力差,且压力越低,水的沸点越低,传热温差越大,物料的水分蒸气压与干燥室内蒸汽分压的压差越大[26],自由水越容易蒸发被抽走,移动性降低。随着干燥的进行,物料表面水分减少,内部扩散阻力增加,水分的不易流动性增强,各状态水分的自由度降低,引起蒸发速度减慢[3]。
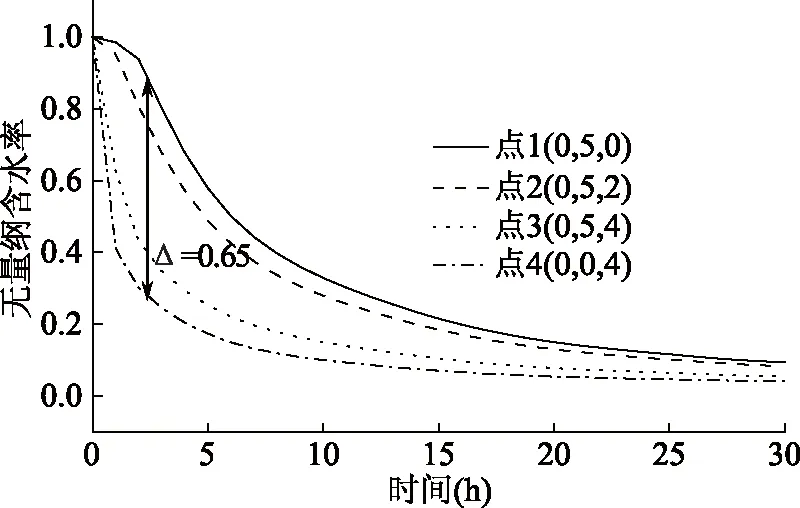
图11 蚕豆各内部点无量纲含水率变化曲线
2.2.5 温度和压力对蚕豆脱水特性的影响 在不同温度和真空条件下,蚕豆脱水过程中无量纲含水率随时间的变化如图12所示。结果表明,在所有情况下,含水率均随温度和真空度的升高而迅速降低。随着干燥腔内压力的降低,即真空度增加,周围空气中的水分浓度降低,分压梯度引起气相水分向表面迁移,从而促进水分蒸发;随着温度升高,水分蒸发加快,蒸发速率增大,加快脱水过程。而在同一时刻,8 ℃脱水环境下的蚕豆含水率比0 ℃时的含水率降低了29.925%;99 kPa脱水环境下的蚕豆含水率比95 kPa时的含水率降低了39.273%。这表明真空压力对含水率影响更大。并且随着温度和真空度的升高,脱水时间也缩短。在97 kPa下,温度从0 ℃上升到8 ℃,脱水时间缩短了7 h。在4 ℃条件下,从95~99 kPa,脱水时间缩短5.5 h。这是由于温度和真空度的升高,提高了水蒸气压力,从而加强了蚕豆内部的水分传递,缩短了脱水时间。此外,温度对脱水时间的影响更为重要。因此在种子保存时,若要得到含水率更低的种子,可以考虑适当增大真空度来实现;而若要快速达到指定含水率状态,可以通过提高脱水温度来控制。

图12 不同环境温度(a)或真空度(b)下含水率变化模拟曲线
3 结论
本文通过实验并模拟的方法研究了不同温度和真空度条件下蚕豆种子的脱水过程,结合核磁共振技术检验了不同时刻种子内部的水分相态,分析了温度、压力对蚕豆脱水特性的影响,研究表明,蚕豆组织有三种水分相态,在自然成熟后摘取的蚕豆种子内部通常会同时存在。自由水游离于细胞间隙,当外界施加压力和温度时,迅速向外排出。而半结合水在自由水大部分被脱除前不会向外逸出,且在脱水前期会有部分自由水和结合水向其转化,导致含量增多。结合水受到大分子束缚,为维持细胞内外水分浓度平衡,含量仅小幅度波动变化,只有长时间脱水才有可能使之完全脱完;真空压力对脱水样品整体的含水率影响更大,而温度对脱水时间的影响更为重要,在种子保存时,应根据期望样品达到的状态以及可接受的脱水时间适当提高真空压力或者脱水温度。
本研究通过研究脱水过程蚕豆种子微观和宏观的水分变化,为种子工业化保存提供了理论依据。而脱水过程中还伴随着种子的收缩变形,未来应着重研究种子力学行为与水分扩散之间的关联以及相互影响,完善种子保存的理论基础。
猜你喜欢
小读者(2021年2期)2021-03-29 05:03:30
能源工程(2020年6期)2021-01-26 00:55:18
公民与法治(2020年13期)2020-07-27 01:30:36
中国奶牛(2019年1期)2019-02-15 07:19:46
学生天地(2018年9期)2018-09-06 13:11:18
现代园艺(2017年11期)2017-06-28 11:32:37
材料科学与工程学报(2016年4期)2017-01-15 13:35:51
高原山地气象研究(2016年3期)2016-02-28 13:53:20
中国塑料(2015年4期)2015-10-14 01:09:21
石油化工应用(2014年11期)2014-03-11 17:40:44
