
发芽处理对青稞β葡聚糖抗氧化和抗炎作用的影响
2020-09-23 12:32孙昌武谢云飞姚卫蓉郭亚辉成玉梁
食品工业科技 2020年17期
孙昌武,谢云飞,姚卫蓉,郭亚辉,成玉梁,钱 和
(江南大学食品学院,江苏无锡 214122)
青稞(Hordeum vulgare Linn. var. nudum Hook. F.)是一种高海拔禾谷类农作物,主要生长在青藏高原地区[1-2],近年来由于其β-葡聚糖含量远高于其他谷物而引起人们的关注。β-葡聚糖是一种水溶性膳食纤维,由β-D吡喃葡萄糖单元组成的线性均多糖[3],由约70%的(1→4)键和30%的(1→3)键组成,由于来源和提取分离方法的不同而导致其分子量不同,一般在20~3100 kDa之间[4-5]。炎症性肠病(inflammatory bowel disease,IBD)是一种多因素疾病[6],该病的发病率在全球均呈持续增加趋势。炎症期间,肠道免疫反应过度亢进引起肠粘膜损伤,同时肠道内部的氧化还原失衡,过氧化物的过量产生会导致肠道进一步的损伤[7]。据文献报道,适量摄入β-葡聚糖对人体肠道有益,它可通过抗氧化特性和抗炎特性来缓解肠道在炎症期间受到的损伤[8-11]。
研究发现,分子量的降低或结构的改变会增强β-葡聚糖的抗氧化能力,改善其抗炎作用[12-14]。已经有采取物理或化学方法如辐射、热处理、酸解来降解多糖[15-17],但很少有关于发芽改变β-葡聚糖分子量从而改变其功能活性的文献。有报道称发芽过程中β-葡聚糖内切酶的活性增加[18],这可能会引起β-葡聚糖的分子量和结构的变化。
本文对青稞进行发芽处理,采用水溶醇沉法并经过耐高温α-淀粉酶、葡萄糖淀粉酶、木聚糖酶和胰蛋白酶除杂提取青稞β-葡聚糖,比较其体外抗氧化能力,并用脂多糖(LPS)诱导肠上皮细胞(Caco-2细胞)的炎症模型来研究β-葡聚糖的抗炎作用,以期为发芽改善青稞β-葡聚糖对炎症期间肠道的保护能力提供理论依据。
1 材料与方法
1.1 材料与仪器
隆子黑青稞 西藏藏缘青稞有限公司提供;人结肠癌细胞Caco-2、完全培养液(内含MEM培养基、20%胎牛血清、1%谷氨酰胺、1%非必需氨基酸和1%丙酮酸钠) 中国科学院上海细胞库;细胞存活率检测试剂盒(CCK-8试剂盒) 上海碧云天生物技术有限公司;肿瘤坏死因子(TNF-α)、白细胞介素-6(IL-6)、前列腺素E2(PGE2)、一氧化氮(NO)、丙二醛(MDA)、超氧化物歧化酶(SOD)、谷胱甘肽(GSH)、FRAP铁离子还原能力测定(FRAP)试剂盒 南京建成生物有限公司;β-葡聚糖检测试剂盒(混联) Megazyme公司;1,1-二苯基-2-三硝基苯肼(DPPH) 美国Sigma公司;多糖标准品Dextran T-2000(Mw2000000)、Dextran T-300(Mw300600)、Dextran T-150(Mw135030)、Dextran T-10(Mw9750)、Dextran T-5(Mw2700)、葡萄糖(Mw180) 中国食品药品检定研究院;无水乙醇等其他化学试剂 国药集团化学试剂有限公司。
Waters 1525型高效液相色谱仪 美国Waters公司;PL2002电子天平 Mettler Toledo仪器有限公司;M5酶标仪 美国Molecular Devices公司;电热恒温水浴锅 常州市朗越仪器制造有限公司;冷冻干燥机 宁波新芝生物科技股份有限公司;旋转蒸发仪 上海亚荣生化仪器厂。
1.2 实验方法
1.2.1 发芽青稞的制备 挑选无虫蛀及霉斑、颗粒饱满的青稞种子,在室温下用0.05%的次氯酸钠溶液浸泡30 min,对种子表面进行灭菌,然后用去离子水漂洗数次,沥干。发芽前,在黑暗的培养箱中将青稞种子在去离子水中浸泡24 h。用去离子水冲洗后,将其平铺在自动恒温(25 ℃)发芽机上,分别于0、6、12、18、24、30 h取样。收集青稞种子在-20 ℃预冷后进行冷冻干燥,之后打粉通过60目筛后在-20 ℃下储存,然后进行下一步的化学分析。
1.2.2 青稞β-葡聚糖的含量检测和提取 根据Gunjan等[19]的方法进行修改。将50 g青稞粉与500 mL 80%(v/v)乙醇在90 ℃下回流2 h。然后在50 ℃下用1000 mL纯水将水溶性β-葡聚糖提取两次,持续2 h。离心后(4000×g,15 min),将提取物合并,并在60 ℃真空下通过旋转蒸发仪浓缩至原始体积的1/3。然后,依次使用热稳定的α-淀粉酶(20 U/mL),β-1,4-木聚糖内切酶(1 U/mL)和胰酶(5 U/mL)去除淀粉、木聚糖和蛋白质。随后,将提取物加入三倍体积的95%(v/v)乙醇在4 ℃下过夜。将沉淀物用70%的乙醇洗涤两次,然后在80 ℃的纯水中重新溶解。离心(4000×g,15 min)后,将上清液转移至3 kDa的超滤管中,通过离心(3500×g,25 min)六次彻底除去分子量小于3 kDa的化合物。最后,将可溶性β-葡聚糖冷冻干燥,得分别发芽0、6、12、18、24、30 h的青稞β-葡聚糖记作QBG0、QBG6、QBG12、QBG18、QBG24、QBG30。使用β-葡聚糖检测试剂盒(混联)检测发芽过程中β-葡聚糖含量和提取后的纯度。
1.2.3 青稞β-葡聚糖的分子量测定 QBG的重均分子量(Mw)和多分散度(Mw/Mn)是通过高效凝胶过滤色谱法检测。将样品配成5 mg/mL的水溶液,用0.22 μm水系微孔过滤后进样20 μL,采用色谱柱UltrahydrogelTMLinear(300 mm×7.8 mm),检测器为Waters 2414示差折光检测器,以0.1 mol/L NaNO3为流动相,流速为0.9 mL/min,柱温45 ℃。
将Dextran T-2000、Dextran T-300、Dextran T-150、Dextran T-10、Dextran T-5、葡萄糖相继进样,记录各自的保留时间TR。得到多糖分子量与保留时间回归方程:logMw=-0.459TR+12.3(R2=0.9952),其中Mw为重均分子量,TR为保留时间。待测样品以相同条件进样,根据回归方程计算得到其重均分子量。
1.2.4 青稞β-葡聚糖的抗氧化能力 将1.2.2得到的青稞β-葡聚糖于去离子水中溶解成浓度为10 mg/mL的多糖溶液作后续抗氧化能力测定。
1.2.4.1 铁离子还原能力的测定(FRAP) FRAP值根据Benzie等[20]的方法测定。将乙酸盐缓冲液(300 mmol/L,pH=3.6)、10 mmol/L的TPTZ盐酸溶液(40 mmol/L)和20 mmol/L FeCl3溶液按照10∶1∶1的比例混合配制FRAP试剂,在37 ℃水浴锅中温浴,现用现配。将0.1 mL多糖溶液(10 mg/mL)与3 mL FRAP试剂混合,室温下反应6 min后立即在593 nm处检测吸光度。用不同浓度的FeSO4·7H2O溶液绘制标准曲线,得到标准曲线公式为Y=7.2048X-0.0103(R2=0.9991)。其中X为吸光度;Y为FRAP值,单位mmol/L Fe2+。
1.2.4.2 DPPH自由基抑制率的测定 根据Kozarski等[21]的方法进行修改来测定。将1 mLβ-葡聚糖溶液(10 mg/mL)与1 mL新鲜制备的0.2 mmol/L DPPH溶液(用纯DMSO配置)以及2 mL DMSO混合,避光放置30 min,以纯DMSO调零测定其在517 nm的吸光度,此为样品组Ai。将每个样品与3 mL DMSO溶液混合,避光放置30 min,以纯DMSO调零测定其在517 nm的吸光度,此为对照组Aj。再测定1 mL DPPH溶液与3 mL纯DSMO混合溶液在517 nm的吸光值,此为空白组的Ac。DPPH自由基抑制率由方程得出:
1.2.4.3 ABTS(2,2′-联氮-双-3-乙基苯并噻唑啉-6-磺酸)自由基抑制率测定 根据Arnao等[22]的方法进行修改来测定。将7.4 mmol/L ABTS溶液和2.6 mmol/L K2SO4溶液等量混合,室温下黑暗中反应12 h制备ABTS工作液。然后通过将1 mL ABTS溶液与60 mL甲醇混合来稀释工作液,使工作液在734 nm吸光度为1.1±0.02。将150 μL多糖溶液(10 mg/mL)与2850 μL稀释后的ABTS工作液在黑暗中反应2 h,以甲醇作为空白在734 nm处检测吸光度。
式中:As为样品吸光度,Ac为蒸馏水代替样品吸光度。
1.2.5 发芽青稞β-葡聚糖对LPS诱导Caco-2细胞炎症模型的影响
1.2.5.1 细胞培养 将Caco-2细胞加入完全培养基复苏后传至第三代,于37 ℃、5% CO2培养箱中培养,每2 d更换一次培养液,待细胞贴壁生长至80%时用胰酶-EDTA按比例消化传代。
1.2.5.2 不同发芽时期的β-葡聚糖对Caco-2细胞存活率的影响 取对生长期细胞,以2×105个/mL的密度在96孔板中均匀加入200 μL,每组5个平行,等细胞达到70%贴壁时,吸掉培养基,先后加入含有β-葡聚糖和LPS的培养基,使β-葡聚糖终浓度达到200 μg/mL,LPS浓度为10 μg/mL,48 h后,每孔加入10 μL CCK-8,孵育2 h后在450 nm下读板。
1.2.5.3 细胞因子和氧化应激参数检测 将细胞用青稞β-葡聚糖和LPS处理并培养48 h后,小心收集细胞加入裂解液后离心取上清液冻存,使用试剂盒确定TNF-α、IL-6、NO、PGE2、SOD、GSH、MDA水平。
1.3 数据处理
利用graphpad prism 6.0软件和SPSS 20.0处理数据。
2 结果与分析
2.1 发芽过程青稞β-葡聚糖含量和分子量变化
由表1可以看出,未发芽青稞β-葡聚糖的含量为3.77%,高于姚豪颖叶[23]得出来的平均值2.45%,青稞β-葡聚糖含量在发芽前24 h含量一直上升,最高含量为4.60%,这与王波等[24]的研究结果有一定差别,原因可能是检测方法差异导致部分不可溶性β-葡聚糖未检出,也有可能是由于品种、品质差异。β-葡聚糖的分子量在发芽期间显著降低(P<0.05),未发芽前β-葡聚糖的重均分子量为275.74 kDa,经过30 h后,分子量降低到114.63 kDa。该结果与Ahmad等[25]研究大麦发芽结果一致,这可能是因为在发芽过程中,存在于谷物种子胚乳中的β-葡聚糖酶活性明显增加,对β-葡聚糖产生了降解作用[26]。
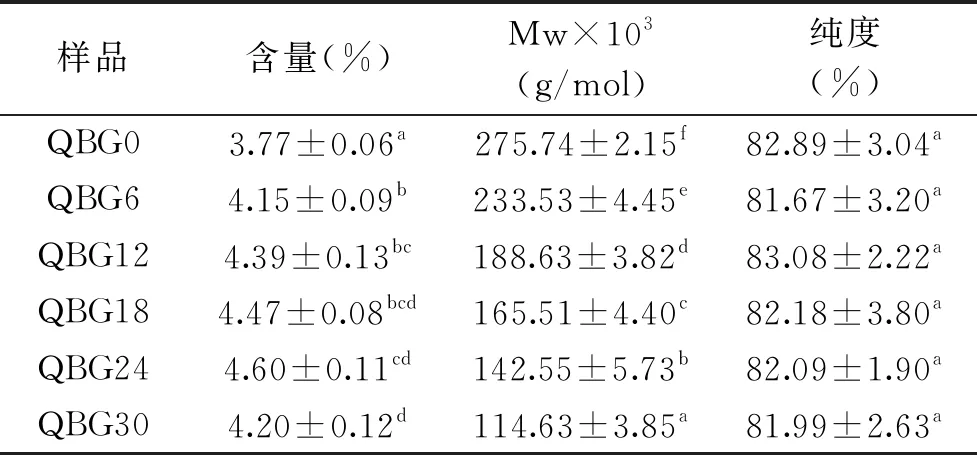
表1 发芽过程青稞β-葡聚糖含量和分子量变化
2.2 发芽过程青稞β-葡聚糖的抗氧化能力变化
图1显示了不同发芽时间青稞β-葡聚糖(10 mg/mL)的抗氧化能力变化。FRAP值反映了样品的还原能力[27]。如图1(A)所示,未发芽青稞中β-葡聚糖的FRAP值为2.47 mmol/L Fe2+,经过发芽处理后,其还原能力持续上升,在30 h达到最高值3.35 mmol/L Fe2+,上升了35.63%,说明发芽使青稞β-葡聚糖的总还原能力得到显著增强(P<0.05)。

图1 发芽过程青稞β-葡聚糖的抗氧化能力变化
DPPH·和ABTS+·是用于确定β-葡聚糖抗氧化活性的自由基,适用于亲水性和亲脂性物质的抗氧化能力测定。如图1(B)可以看出,未发芽的青稞β-葡聚糖的DPPH自由基抑制率为5.53%,随着发芽时间的延长,在30 h时其值达到最大值19.28%,ABTS自由基抑制率变化趋势与之相似,发芽30 h从9.11%提升到16.45%。
尽管这三种分析方法测定抗氧化能力的机理并不完全相同,但上述实验结果一致表明发芽可以显著(P<0.05)提高青稞β-葡聚糖的抗氧化能力。多糖的分子量与其抗氧化能力具有直接关系,如潘莹等[28]、徐雪峰[29]等均发现分子量不同的植物多糖组分对自由基的清除能力不同。发芽过程中β-葡聚糖酶的作用增强,导致青稞中β-葡聚糖含量发生变化,而在同等浓度下,发芽时间越长,获得的青稞β-葡聚糖分子量越低,可能使羟基暴露或者产生了更多可以充当自由基猝灭剂的异头氢,使其抗氧化能力得到增强[30]。从24到30 hβ-葡聚糖含量下降不显著(P>0.05),但其抗氧化能力上升显著(P<0.05),说明经过发芽处理后的青稞β-葡聚糖更加具有作为抗氧化剂的潜力。
2.3 发芽青稞β-葡聚糖对LPS诱导Caco-2细胞炎症模型的影响
2.3.1 细胞存活率 如图2模型组(LPS组)所示,Caco-2细胞经过LPS(10 μg/mL)处理后,细胞的存活率较空白组(CON组)相比下降了51.25%。当加入不同发芽时间的β-葡聚糖(200 μg/mL)时,Caco-2细胞的存活率较LPS组有所上升,且发芽时间越长,得到的β-葡聚糖对抑制LPS诱导的Caco-2细胞炎症模型效果越明显,发芽时间30 h提取的β-葡聚糖对细胞的保护作用最强,存活率最高达到69.27%。

图2 发芽青稞β-葡聚糖对Caco-2细胞炎症期间存活率的影响
2.3.2 细胞因子 通过测定炎症因子来评估β-葡聚糖对LPS诱导的Caco-2细胞炎症状态的影响。TNF-α是参与IBD病程中组织损伤的关键促炎免疫介质,其含量的下降与IBD的改善密切相关[31-32]。IL-6是炎症反应中重要的细胞因子,在机体发生炎症时,细胞分泌IL-6可以促进炎症反应。如图3(A)所示,TNF-α在空白组细胞中的含量为139.75 ng/L,当受到10 μg/mL LPS刺激时,TNF-α的含量显著提升到462.19 ng/L(P<0.05),当加入200 μg/mL发芽青稞β-葡聚糖干预时,其含量得到显著抑制(P<0.05),青稞发芽时间越长获得的β-葡聚糖对炎症期间Caco-2细胞分泌的TNF-α抑制越强,抑制能力最强为发芽30 h的青稞β-葡聚糖,TNF-α含量降低到了274.87 ng/L,与未发芽青稞获得的β-葡聚糖相比降低了40.53%。图3(B)中IL-6的含量也是在模型组中显著上升(P<0.05),样品组中发芽青稞β-葡聚糖能够显著减少Caco-2细胞分泌的IL-6(P<0.05),最高降低到58.15%。这说明发芽青稞β-葡聚糖能够显著降低Caco-2细胞在LPS诱导的炎症模型中分泌的TNF-α和IL-6含量,对于调节炎症具有至关重要的作用。

图3 青稞β-葡聚糖对Caco-2细胞炎症期间细胞因子分泌的影响
促炎因子介导机体体液免疫和细胞免疫,促进炎症反应以达到快速清除抗原的目的。然而,过度表达的促炎症因子会导致炎症反应失控,持续的免疫应答将会导致机体的自我破坏,如PGE2、NO的显著升高可以诱发氧化损伤造成正常细胞和组织的炎症反应[33]。因此,在炎症后期抑制促炎症因子的表达同时促进抗炎因子的表达有助于恢复机体免疫功能平衡。如图3所示,与正常对照组相比,LPS刺激能显著诱导Caco-2细胞促炎因子PGE2和NO的释放(P<0.05),而与模型组相比,200 μg/mL的β-葡聚糖能使损伤细胞中的PGE2、NO分泌水平分别下降48.84%、32.47%,反映β-葡聚糖可抑制LPS诱导的肠上皮细胞炎症反应,且发芽时间越长得到的青稞β-葡聚糖效果越明显。由于发芽会降低青稞β-葡聚糖的分子量,因此可以推断出分子量越小,其抗炎作用越强,与Lin等[34]的研究一致,其原因可能是分子量越小,β-葡聚糖的低粘度和高流动性使其能够更好地与Caco-2细胞表面受体接触并发挥其抗炎作用。
2.3.3 氧化应激参数 在肠炎期间,由于机体内的过度免疫反应会产生过量的活性氧(reactive oxygen species,ROS),导致氧化还原水平失衡,从而使肠道受到一定的氧化损伤。具有抗氧化活性的物质能够中和或转移ROS,例如超氧化物歧化酶(SOD)、谷胱甘肽(GSH)等可以减轻IBD期间的肠道损伤,改善肠上皮细胞的屏障功能[35]。如图4(B)、4(C)所示该实验中通过β-葡聚糖的干预可以显著(P<0.05)增加炎症期间SOD的活性和GSH的分泌水平来维持细胞内的氧化还原的动态平衡,减少自由基对自身的损害作用。丙二醛(MDA)则是分子物质发生脂质过氧化反应而形成的一种代表性脂质过氧化降解产物[36],能够间接反映出细胞或组织受到氧自由基攻击后受损程度。如图4(A)所示,经过发芽后的β-葡聚糖能够显著(P<0.05)降低MDA在炎症期间的产生,从而改善细胞在炎症期间受到的氧化损伤。综上所述,小分子量的β-葡聚糖可以显著改善肠道炎症期间氧化应激水平,从而缓解肠道细胞受到二次损伤。

图4 青稞β-葡聚糖对Caco-2细胞炎症期间氧化应激参数的影响
3 结论
本文主要研究了发芽过程中青稞β-葡聚糖通过抗氧化能力和抗炎作用对炎症期间肠道的保护作用。研究结果显示,青稞β-葡聚糖的含量在发芽过程中总体呈现先上升后下降的趋势,分子量在发芽过程中呈现下降趋势;同时,发芽30 h提取的β-葡聚糖呈现出更好的抗氧化性且能够有效地抑制LPS造成的Caco-2细胞炎症损伤,提高细胞存活率,显著抑制炎症模型中细胞因子TNF-α、IL-6、PGE2、NO的分泌,并且有效改善细胞炎症期间的氧化应激水平。综上所述,发芽会降低青稞β-葡聚糖分子量并增强其抗氧化能力和抗炎作用,从而缓解肠道在炎症期间所受到的损伤,因此在食品工业中具有更好的发展潜力。而发芽导致青稞β-葡聚糖的分子量下降,其结构是否发生变化,如何建立青稞β-葡聚糖的分子量、结构与生物活性之间的构效关系也是今后研究的方向。
猜你喜欢
石油沥青(2022年2期)2022-05-23
星星·诗歌原创(2022年1期)2022-04-16
乡村科技(2021年12期)2021-09-06
纺织科学研究(2021年7期)2021-08-14
西藏人文地理(2020年4期)2020-11-19
翠苑(2019年4期)2019-11-11
中国糖料(2019年1期)2019-02-13
中国酿造(2018年10期)2018-11-03
中国野生植物资源(2018年3期)2018-08-06
农业环境科学学报(2017年2期)2017-03-20
