
濒危植物千果榄仁种群和群落学研究
2020-09-23 08:20李汶霏邓莉兰
湖北民族大学学报(自然科学版) 2020年3期
余 潇,张 宝,李汶霏,邓莉兰
(1.西南林业大学 园林园艺学院,昆明 650224;2.西南林业大学 林学院,昆明 650224;3.西南山地森林资源保育与利用教育部重点实验室,昆明 650224)
千果榄仁(Terminaliamyriocapra)为使君子科(Combretaceae)诃子属 (Terminalia)常绿或半常绿乔木[1],是北亚热带野生植物资源的重要组成部分,为我国二级重点保护野生植物,按IUCN地方濒危等级标准评价属于“濒危种(EN)”.谢春华等[2-3]对千果榄仁的地理分布研究表明,千果榄仁在我国主要分布广西(龙津)、云南(中部至南部)和西藏(墨脱),为产区的习见上层树种之一,是我国热带雨林和季雨林的代表种和指示种.目前,国内外对千果榄仁各方面的研究比较少,研究主要集中在千果榄仁的形态特征[2]、生物学特性[2]、化学药用价值[4]、遗传育种[5-6]等方面.本研究以铜壁关自然保护区千果榄仁典型样地群落进行调查分析,有利于填补该物种群落生态研究的空白,对研究热带雨林和季雨林的更新、演替和生态评价等具有重要意义[7],也为千果榄仁合理保护及人工恢复以提供科学依据.
1 研究区概况
研究区位于云南省西南部德宏州瑞丽市,处于东经97°31′~98°02′,北纬23°38′~24°14′之间,地处横断山脉高黎贡山余脉的向南延伸部分,地势西北高东南低.研究区境内的云南省铜壁关省级自然保护区为我国40处A级(具有全球保护意义)保护区之一,是唯一分布在我国境内伊洛瓦底江流域热带区域的自然保护区,形成了以阿萨姆娑罗双、盈江龙脑香等标志性热带雨林植被.
2 研究方法
2.1 资料收集与确定分布点
通过查阅与收集文献资料、访问植物专家学者、基层访问等方式,根据以往调查所得千果榄仁具体分布位置的基础上,初步获得千果榄仁的分布区范围,并且于地形图上标记出来.
2.2 野外实地调查
调查时间为2015年10月和2016年8月,采用样方法,根据实地调查与勘测,选取了瑞丽市勐卯镇的莫里瀑布风景区以及弄岛镇等戛村的铜壁关自然保护区4个典型样地群落设置20 m×20 m样方,记录样方内目的物种的数量、海拔、树高、胸径、冠幅、坡向、坡位、高度、人为影响等.再将样地划分成16个5 m×5 m的小样方记录调查乔木层,对于样方内胸径大于5 cm的乔木逐一记录,记录种名、株数、株高、胸径、冠幅;样方的4个边角及中心点各设置1个5 m×5 m的灌木样方和2 m×2 m的草本样方,记录灌木和草本的种名、密度、高度、盖度等[8].
2.3 数据分析
1)种群径级结构的分析.由于乔木其个别树龄无法确定,因此实地调查参考前人研究以其胸径对树龄进行推测[9-10].根据谢春华等[2]的观察研究,千果榄仁年均胸径生长量可达2.0 cm,成年树胸径一般可达1 m以上[11].径级结构图的绘制以2 cm为一个间隔,表示千果榄仁群落的动态发展.千果榄仁一般在种植后14年开始开花结果[2],所以径级结构分析取28 cm作为幼树和成年树的临界值.以此为依据统计各龄级千果榄仁的个体数量.
2)种群的高度级结构分析.根据野外调查的数据,结合谢春华等[2]的观察研究,千果榄仁成年树年均树高生长量分别可达1.50 m.将记录到的千果榄仁种群高度级以1.5 m为一个高度级进行划分,共划分为24个高度级[11].与径级结构分析一样,取植株种植14年后的高度21 m为幼树和成年树的临界值.
3)生活型和叶级性质进行分析.根据生活型系统的分类标准[9-10],将高等植物划分为高位芽、地上芽、地面芽、地下芽植物和一年生植物5大类,将其生活型进行归类统计.根据丹麦生态学家Raunkiaer C对群落中植物个体叶片的划分方法,将对叶片的描述分为叶级、叶型、叶质和叶缘4个方面[9-10],对它们进行归类统计.
3 结果与分析
3.1 千果榄仁群落种群学分析
3.1.1 生境特征及现状 从生境上来看,千果榄仁主要分布于海拔在800~1 200 m的峭壁岩石附近的中下坡位置或平地上,坡向多数为较陡的阳坡,且以大乔木形态散点分布,这与文献资料所记载的千果榄仁生活习性类似.千果榄仁所处的群落属于季节雨林,由于所处群落的缘故,所有群落的郁闭度和盖度均较高.千果榄仁分布区内4个样地群落的土壤类型均为赤红壤和黄壤,土壤pH值在5.5~6.0之间,属于酸性土壤.由于分布点位于风景区内及道路旁,受到旅游开发、放牧采集等人为影响较大,导致千果榄仁的数量锐减以及生境恶化.
3.1.2 种群径级结构分析 由表1可知,千果榄仁群落中以胸径≥40 m的植株分布最多,共6株,占总株数的40%;而胸径≤28 cm的植株共6株,占总株数的40%.群落中最小的一株高3 m,胸径10 cm;最大的一株高35 m,胸径50 cm.从数据统计分析中可以看出,胸径≤28 cm的幼树个体与成年高大乔木胸径≥40 cm的数量大致相同.其径级结构图如图1所示,可以看出千果榄仁种群树龄构造表现出显著的的倒三角形,幼苗个体稀少,胸径在40 cm以上的成年大树较多,分析可知种群径级结构为衰退型.
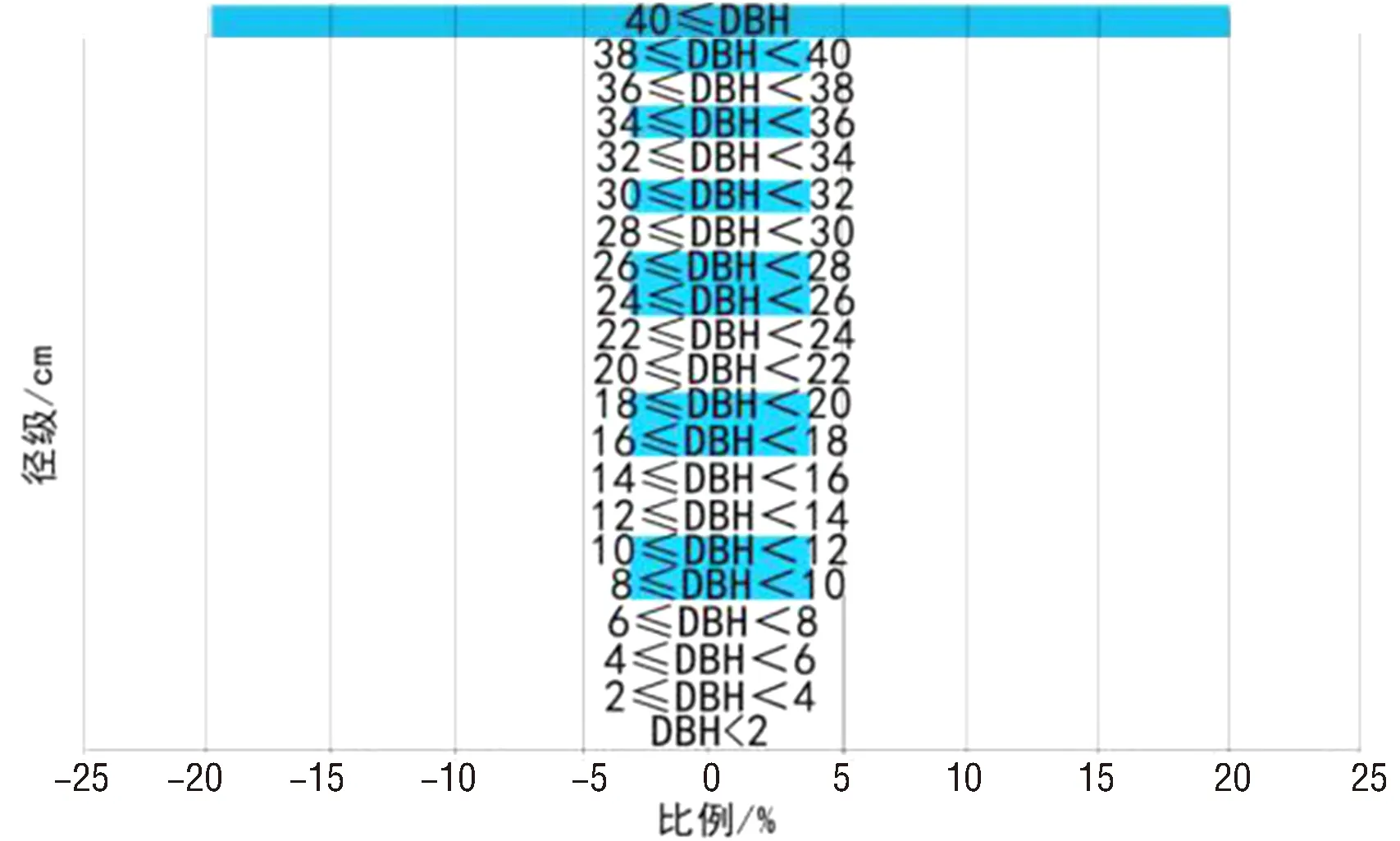
图1 千果榄仁径级结构Fig.1 The diameter-class structure of T.myriocarpa population
表1 野生千果榄仁径级结构
Tab.1 The diameter-class structure ofT.myriocarpapopulation

径级/cm株数比例/%径级/cm株数比例/%径级/cm株数比例/%DBH<200.0014≤DBH<1600.0028≤DBH<3000.002≤DBH<400.0016≤DBH<1816.6730≤DBH<3216.674≤DBH<600.0018≤DBH<2016.6732≤DBH<3400.006≤DBH<800.0020≤DBH<2200.0034≤DBH<3616.678≤DBH<1016.6722≤DBH<2400.0036≤DBH<3800.0010≤DBH<1216.6724≤DBH<2616.6738≤DBH<4016.6712≤DBH<1400.0026≤DBH<2816.6740≤DBH640.00
3.1.3 种群高度级结构分析 根据表2可知,千果榄仁种群的高度级结构波动较小,幼树个体数量与成年个体大致相当,数量相似,种群结构相对平衡.但千果榄仁的高度和胸径增长确并没有表现出一致性,这说明千果榄仁为典型的喜阳中高位植物乔木,为热带季雨林速生树种,幼树生长主要表现在高度级结构的变化,胸径的增长量滞后于高度的生长量;而成年大树高度增量速度减慢,胸径增长量加快.

表2 千果榄仁高度级结构Tab.2 The high class structure of T.myriocarpa population
3.1.4 种群结构及更新 天然环境下,千果榄仁的繁殖主要包括以种子繁殖为主,但千果榄仁种子小(2~3 mm),饱实率极低,所调查的群落观察到千果榄仁都为高大乔木,各群落内的幼苗、幼树少,导致了千果榄仁种群的更新困难.千果榄仁群落径级结构表现出显著的的倒金字塔形,可知该地的千果榄仁由于种群更新较差,处于衰退阶段.根据现场调查的数据并结合相关研究资料显示,千果榄仁的种子发芽率较低,种子寿命较短,发芽力弱,常温储藏4个月后种子即丧失发芽力,在自然环境中的发芽率不足20%.通过结合分析可以得出千果榄仁天然更新能力差,自然增殖能力弱,在荫蔽的热带雨林内几乎不见幼树,仅在向阳和空旷的地方偶见幼苗生长情况[12].
究其更新不良的原因.一方面由于千果榄仁群落遭受较大的人为干扰影响,生境破坏较严重且同时期生长环境恶劣,人为活动的强烈干扰,如采集、放牧、偷伐等严重,导致次生物种大量进入,原生物种逐渐消失,次生度增大,使得适合千果榄仁生存的生境逐渐减少,幼苗变少,种群更新困难,物种面临灭绝的挑战.另一方面是由于千果榄仁的野生资源散生、结实率低且在自然环境中发芽率低,果实常又遭牲畜的啃食.
3.2 千果榄仁群落学分析
3.2.1 群落物种组成 根据外业调查的数据进行内业整理计算,4个调查样地共计1 600 m2的群落内共记录维管束植物67种,隶属于44科65属.其中蕨类植物6科6属6种,如金毛狗、贯众、观音莲座蕨等.裸子植物1科1属1种,为直立买麻藤;被子植物37科58属60种,其中双子叶植物30科48属50种,如纽子果、红光树、西南木荷等;单子叶植物7科10属10种,如鱼尾葵、野龙竹、杜若等.乔木种类27种,灌木种类12种,草本28种,乔灌草所占比例为4∶1.8∶4.2.
由于瑞丽市独特的地理区位,本地区为北回归线以北的低海拔区域,气候上广受印度洋暖湿气流影响,温度常年保持在15~20℃的水平,热带东南亚的气候特征明显,在植物相关性上与缅甸和印度联系密切[13-18].根据统计,千果榄仁群落丰富度较高,常绿阔叶植被是主要类型.从群落的结构层次来分析,乔木层、灌木层及草本层的物种分布量分别是26种、13种和28种.所占群落内物种比例分别是40.30%、19.40%和40.30%,可以看出乔灌草各层的比例相对合理.群落内物种有一部分是以同科的形式出现,其中最多的属是大戟科(Euphorbiaceae)和菊科(Compositae)的植物,各有4种,其次豆科(Leguminosae)、樟科(Lauraceae)、茜草科(Rubiaceae)和禾本科(Gramineae),各有3种.其余的物种多以每科2种及单科单属的形式存在,如爵床、线柱苣苔、八宝树等.
3.2.2 群落种子植物区系分析 根据植物区系的区划分析,本区地处东南亚(印度—马来西亚)热带生物区系向东亚亚热带—温带生物区系的过渡地带,主要属于热带性质的植物区系,其所具有的明显特征即为热带亚洲植物区系的特征,然而在热带亚洲成分中,印度(喜马拉雅)-缅甸植物区系是其受到影响最为强烈的区系[15-27].区系分析参考吴征镒的《世界种子植物区系》关于种子植物属的区系划分原则,该物种记录的种子植物区系地理如表3所示.
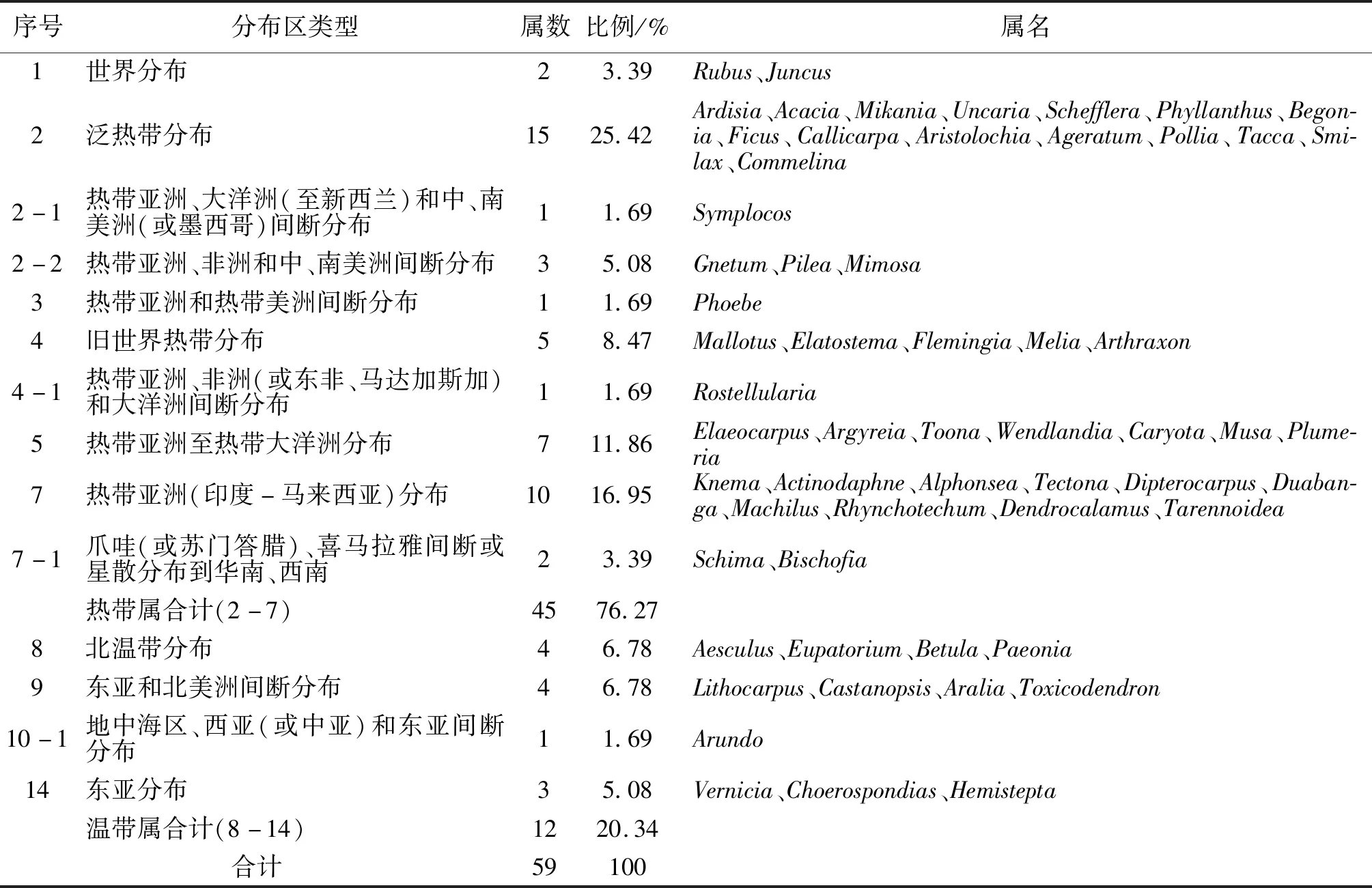
表3 瑞丽市千果榄仁群落种子植物区系统计Tab.3 The generic areal-types of T.myriocarpa community in Ruili County
整个群落内共计种子植物59个属,属的地理成分分布在10大类型内,共计14个小类型.参考《中国种子植物属的分布区类型》属的地理成分分类[28],该地区属的分布大类占到了总数的约56%,可见其植被种类的丰富程度.由表3可知,千果榄仁群落内热带分布型最多,达45属,占总属数的76.27%,又以泛热带分布为主,有15属,占总属数的25.42%;温带分布型属共12属,占总属数的20.34%,其中北温带分布、东亚和北美洲间断分布均分布4种,各占总属数的6.78%,东亚分布型共3属,占总数的5.08%.该地区千果榄仁群落的植物区系以热带分布为主要分布型,这与我国泛滇老缅越地区植物区系中以泛热带分布为主的现状相同,反映了瑞丽地区的热带植物群落性质.
泛热带分布、热带亚洲至热带大洋洲分布和热带亚洲(印度-马来西亚)分布共计32属,占总属数的54.24%,如Ardisia、Acacia、Elaeocarpus、Argyreia、Knema、Actinodaphne等,表明千果榄仁群落植物区系的热带亚洲亲缘性,与热带大洋洲植物区系有一定的联系,也体现了该区域是热带亚洲的北延部分的气候性质.瑞丽市位于云南省西南部,中南半岛北缘,距离北回归线较近,为我国较北的热带雨林地区,瑞丽市拥有特殊的气候,终年温度较高,在地形上属于青藏高原-云贵高原向平原地区的过度地带,其拥有独特的土壤和湿度条件[29].由于该地带是一个过渡地带,为东南亚热带生物区系向东亚亚热带-温带生物区系之间的一个缓冲过渡区域,地理条件较为特殊,造成了它的生物多样性较为丰富,这也就解释了其热带成分占比较大的原因.它们中有很多是热带地区森林群落中的恒有成分,如榕属、杜英属、鹅掌柴属等,占据着热带森林群落的各个层片.
所有的温带性质的属有12属,占总属数的20.34%,远远低于我国同类属的比例(40.87%),但明显高于云南南部国家级自然保护区温带属的比例(15.55%)[12-16].这是因为,就全国而言,这里纬度偏南,海拔低,受印度洋孟加拉湾暖湿气流的影响,海拔700 m以下属北热带气候类型,植物区系的温带成分较国内其他地区为低;但与同属北热带的其他区域如南滚河国家级自然保护区和西双版纳自然保护区相比较,其纬度与海拔又较高,区内的气候与生境类型更复杂多样化.
3.2.3 群落生活型和叶片性质分析
1)生活型.根据生活型系统的分类标准[8-9]对群落内的目的物种的生活型统计分析.根据表4可知,高位芽植物为千果榄仁群落中的主要生活型,占总数的65.67%,其中小高位芽植物和矮高位芽植物分别占总数的22.39%和26.87%,说明群落的乔木、小乔木、灌木较丰富.大高位芽仅占2.99%,如大叶风吹楠、鸡蛋花等.由于该地区属于纬度相对较低(北纬23°),海拔较低(平均海拔500 m),热区植物性质明显,促使当地植物的高位芽植物占据主导地位,不同类型相间交错镶嵌,表现出以龙脑香科树种为标志的热带雨林的各种群落成分,因此,千果榄仁群落目的物种的组成以小高位芽植物和矮高位芽植物为主,一年生植物及地下芽植物数量较少,热带雨林和季雨林中高大乔木较多,霜冻少,气温高,独具特色的自然地理环境造就了以之相对应的植物生活型谱.
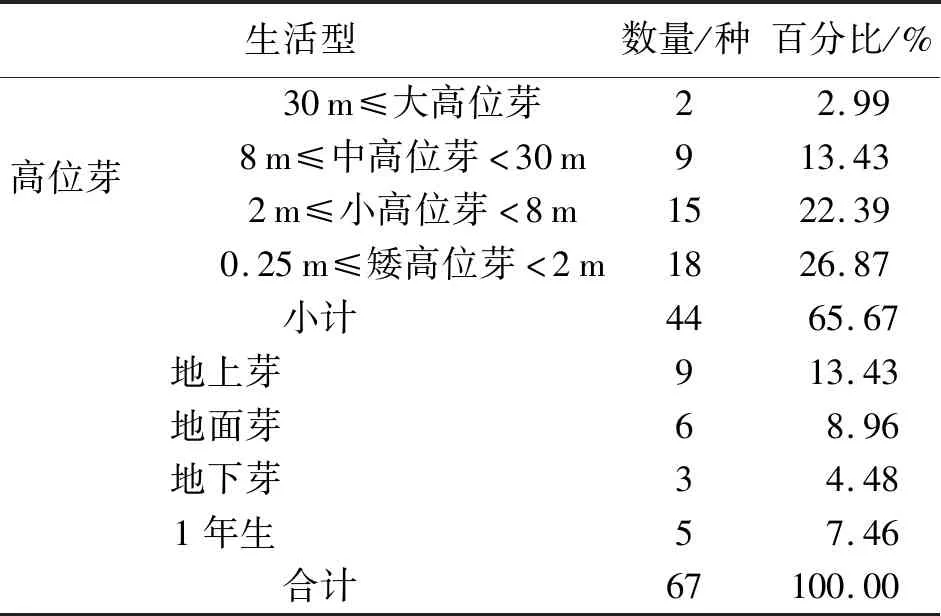
表4 瑞丽市千果榄仁群落生活型统计表Tab.4 The life form statistics of T.myriocarpa community in RuiLi County
2)叶片性质.对千果榄仁群落中的67种维管束植物进行叶片性质统计,统计数据如统计表5所示.千果榄仁群落中,中型叶植物有33种,占比最大,占总数的49.25%,小型叶共计25种,大型叶和巨型叶分别有4种和1种,显示整个群落的叶级以中型叶为主.整个群落以单叶物种为主,达到40种,占群落叶型物种总数的59.70%;叶质以草质为主,叶缘以全缘物种居多.分析可知,群落的叶片性质与该区域的气候相一致,表现为热带地区及泛热带区系气候湿润多雨,植被茂密,植物叶片相较于温带地区树种叶级以中型(2 016~18 225 mm2)为主,叶质以草质为主的特性.

表5 千果榄仁群落植物叶片性质Tab.5 The leaf properties in T.myriocarpa community
3.2.4 群落结构分析
1)群落垂直结构.千果榄仁群落形成垂直层级现象十分明显,根据生长型,千果榄仁群落片层结构可以划分为5个层片.
①大乔木层片:该区域的乔木层较为明显,主要以大叶风吹楠、千果榄仁、云南七叶树等为上层高大乔木树种,上层高大乔木的高度均为20 m及以上,上层乔木枝叶繁茂阻挡了大部分下层植物生长的阳光.该层片为群落的建群层片.②小乔木层片:有的样地上层乔木高度及冠幅盖度相对较低,形成了次一级的小乔木层次,如楤木、藤春、金合欢等.该层片成为建群层片的共建层片或者附属层片.③灌木层片:群落的灌木层植物主要由一些耐荫性较强的树种组成,平均高度在3.5 m左右,以野芭蕉、野龙竹、金毛狗等为主.由于热带雨林、季雨林上层植物的竞争优势,阳光大部分被吸收阻挡,其群落中统计的灌木总数并不如乔木层丰富.该层片在各类群落中都能成为建群层片或共建层片.④多年生高位草本层:群落中统计的草本层共计27种,平均高度16.85 cm,以林下禾本科、菊科植物以及蕨类植物为主,多年生高位草本层主要以蕨类植物如贯众、观音莲座蕨等以及箭根薯为主,是群落的附属层片.⑤一年生或多年生小丛生禾草层片:主要由灯心草、荩草、泥胡菜、薇甘菊等组成,它是热带季雨林群落的共建层片或附属层片.
整个样地群落中,乔木层的种类及数量最多,草本层次之,乔木层的物种量具有绝对优势,高度的平均值为14 m,说明以乔木为竞争优势的热带雨林、季雨林群落是千果榄仁群落的典型垂直结构.
2)群落的水平结构.群落的水平结构是由于在光照、水分以及海拔高度的变化而形成的在水平分布和配置上的差异性.千果榄仁群落的乔木层郁闭度都在0.7~0.9左右,除大叶风吹楠、云南七叶树、千果榄仁等作为优势种分布较为均匀外,很多的非优势种呈零星分布态势,例如西南木荷、红梗润楠、灰木等,灌木和草本在群落内部分布量较少,这与群落位于热带雨林中灌木层和草本层在生存上处于吸收阳光和养分的劣势相一致.灌木植物集群状况:鱼尾葵、鹅掌柴呈小丛、小斑块或大丛状分布;而野芭蕉、野龙竹则呈条带状或团块状集聚分布;其余灌木大部分呈小丛、小斑块或大丛状分布.而草本植物集群方面,除含羞草、野牡丹在群落中呈单生状态分布;其它草本植物如楼梯草、薇甘菊、杜若等都呈大丛或大斑块状分布.
3.2.5 群落重要值分析 从千果榄仁群落乔木层的重要值(如表6所示)分析可知:除千果榄仁外,重要值较高的依次是大叶风吹楠、大叶杜英、云南七叶树、滇南杜英、刺栲、红光树、西南桦、红椿.在灌木层方面(如表7所示),重要值较高的是:野龙竹、鱼尾葵、鹅掌柴、野桐、小柚木、紫珠.因此,从群落重要值方面分析,该群落是以大叶风吹楠、云南七叶树、大叶杜英和野龙竹等主要种群组成的热带季雨林群落,计算结果的分析也与该群落区系较为相似.重要值的大小代表了物种在群落中的生态位,物种的重要值越大,其在群落中的优势就越大,可以当作建群种和优势种.因此在千果榄仁的野外保护和引种时,也应该注意其建群种的栽植,有利于形成稳定的生态群落.

表6 千果榄仁群落乔木层物种重要值Tab.6 Importance values of species in tree layer of T.myriocarpa
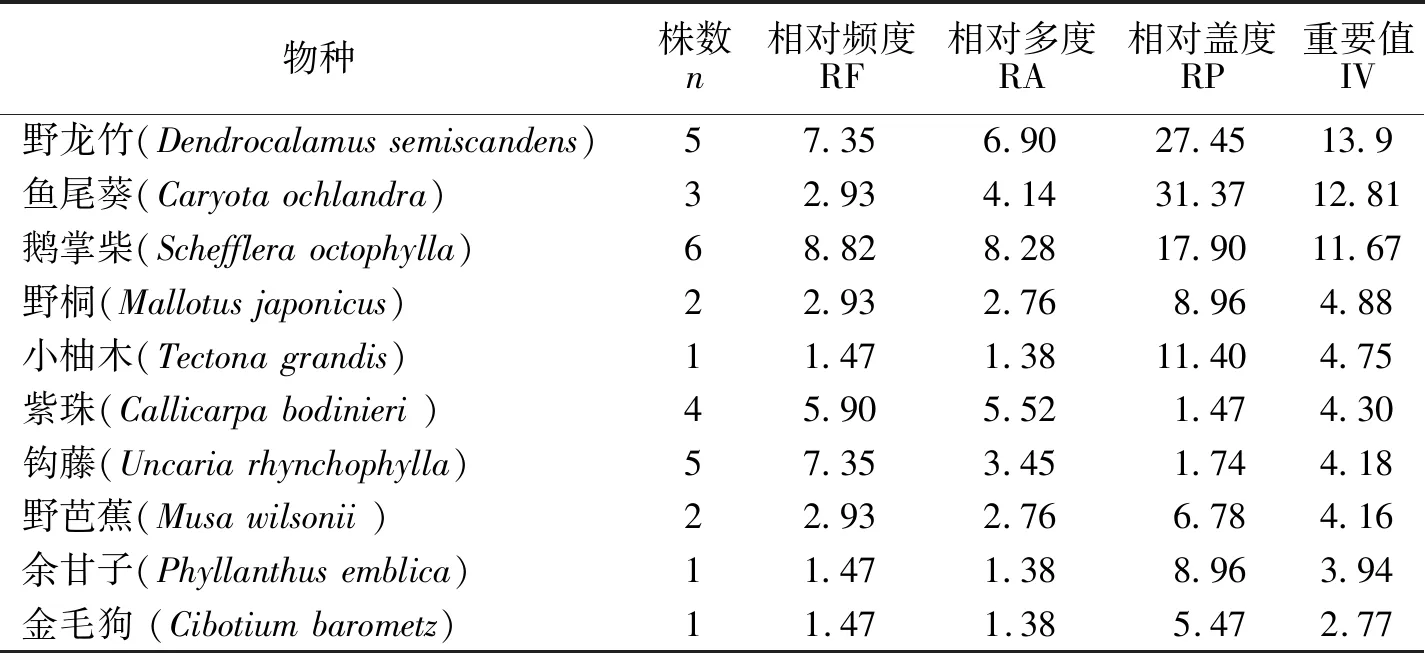
表7 瑞丽市千果榄群落仁灌木层物种重要值Tab.7 Importance values of species in shrub layer of T.myriocarpa in Ruili country
4 讨论
通过数据的分析,得出该地区植物区系有如下几个特征:①具有强烈的亚洲热带性质,其中热带亚热带分布的类型是该区域内数量最多的地理成分.其主要原因是由于该地区立体气候明显,植物的垂直带谱丰富.②具有明显的西南部次大陆特征,该地区千果榄仁群落无论是热带植物组成还是温带植物组成,都显示出其强烈的西南部次大陆特征,其温带植物组成,多数生长于较高海拔段,与喜马拉雅有密切联系;而热带植物组成,则与南亚次大陆有密切联系.这种植物区系的显著西南部次大陆特征,在我国其它地方极少出现此类现象[14-21].③具有一定的热带北缘性质.由于滇西南瑞丽地区热带雨林分布在东南亚热带的北缘,属于我国分布最为北面拥有热带雨林的地区.在云南省,也是分布区在最北面的热带雨林地区,植物区系明显的热带北缘性质被凸显,在属的分布上,由于这一原因,也具有了一些温带成分的出现,这一现象也较好的解释了瑞丽地区的千果榄仁群落中植物区系具有较为明显的热带北缘的性质.该区在热带区系的关系上主要和热带亚洲成分和泛热带联系最为密切;在温带区系的关系上主要和东亚成分特别是东亚和北美洲间断分布联系最为密切.然而,瑞丽市地处云南省西南部,在云南省内,本区与南亚次大陆的联系最为紧密,尤其是和滇老缅边境结合区的相关性最强;而相对于全国而言,本区与喜马拉雅间断分布较为密切[22-29].
千果榄仁在干扰小的情况下,可缓慢恢复种群数量的增长和群落的构建.种群的壮大是一个需要长期演替的过程,千果榄仁增长速度较缓慢,若未来干扰强度变大且缺乏保护,种群将面临较高的死亡风险,甚至消失.因此,要保证千果榄仁种群的稳定发展和生态平衡,首先是了解其生物学特性及生殖繁育机制,促进母树结实,提高种子萌发率;其次是适当进行人工抚育,保证幼苗成活率,使幼苗继续稳定健康生长;最后是加强对其原生境的保护,减少人为破坏、牲畜践踏、生境流失等.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
园艺与种苗(2021年3期)2021-05-26
农村实用技术(2020年1期)2020-12-16
老年教育(2020年12期)2020-12-11
种子(2019年6期)2019-07-13
世界热带农业信息(2019年11期)2019-01-05
读写算·小学中年级版(2017年4期)2017-05-08
湖北农业科学(2014年4期)2014-05-04
学与玩(2009年2期)2009-03-09
