
大气CO2浓度和气温升高对谷子生长及产量的影响
2020-09-01 13:54任宏芳李阿立郝兴宇宗毓铮张东升李萍
生态环境学报 2020年6期
任宏芳,李阿立,郝兴宇,宗毓铮,张东升,李萍
山西农业大学农学院,山西 太谷 030801
随着经济和工农业的飞速发展,大气CO2浓度已经增加到 406 μmol∙mol-1,比工业革命前的 280 μmol∙mol-1增加了 41%(NOAA,2014),预计到2050 年将达 550 μmol∙mol-1,到本世纪末最高或将增加至 936 μmol∙mol-1(IPCC,2013)。CO2是主要的温室气体,CO2浓度增加会引起全球气温升高(秦大河,2007)。在过去的 100年间,大气温度平均升高了0.78 ℃,预计到2050年将继续上升0.8—3.2 ℃(IPCC,2013)。全球变暖和CO2浓度升高会影响作物的生长发育(房世波等,2010)。大气CO2浓度升高会促进作物的光合作用,有利于作物生物量和产量的提高(Leakey et al.,2006;李靖涛等,2015;郝兴宇等,2011)。温度升高对作物发育和生理生化过程也会产生一系列重要影响(张旭等,2002;Peng et al.,2004),同时增温和升高CO2浓度能改变植物能量代谢和物质转化等生理过程,影响植物的生长发育(Morison et al.,1999)。张治安等(2006)研究发现不同CO2和温度水平下人参叶片光合速率的响应特性不同;陈楠楠等(2013)发现大气 CO2浓度升高会导致水稻和小麦的产量增加,但是温度升高会削弱大气CO2浓度升高的增产效应;Gesch et al.(2003)研究发现,CO2浓度升高条件下,温度升高对植物叶片光合速率有抑制作用。苏营等(2016)对大豆(Glycine max(Linn.) Merr.)的观测也表明,CO2浓度升高对大豆株高和茎直径的生长具有促进作用,增温使这种促进作用更大。
谷子(Setaria italica)禾本科狗尾属,中国耐旱稳产作物,北方地区重要的粮食作物之一,不但营养丰富而且具有抗旱性、耐瘠薄和抗病虫等特性,是改善人民膳食结构的重要粮食作物之一(薛月圆等,2008)。有研究表明,CO2浓度升高后,谷子净光合速率提高,株高、茎直径、叶面积增加,叶片叶绿素含量下降(郝兴宇等,2010)。但迄今为止,关于CO2倍增和气温升高交互作用条件下谷子光合生长及产量的研究鲜有报道。本研究拟利用CO2和温度控制气室对谷子影响进行相关研究,了解未来气候变化背景下,大气CO2浓度升高和温度升高对谷子光合生理及生长发育的影响,以更加清楚未来谷子的生产情况,为改进谷子管理栽培技术、提高谷子产量提供理论依据。
1 材料与方法
1.1 试验地点和试供土壤
实验地位于山西省晋中市太谷县山西农业大学农学院实验基地(37.42°N,112.58°E)。供试土壤为褐潮土,播前有机质质量分数2.37%,全N质量分数 1.12 g∙kg-1,速效 N 45.28 mg∙kg-1,速效 P 25.65 mg∙kg-1,速效 K 280.5 mg∙kg-1,装桶前过筛并混匀。试验品种为对晋谷21 EMS诱变后的超早熟品种Xiaomi,该品种株高约40—60 cm,全生育期70 d左右。
1.2 设施构成与试验设计
控制气室结构为钢结构,外罩钢化玻璃,自然采光,透光率80%左右,每个气室面积为4×8 m2,高3.2 m。共4个室,每个气室面积一致。采用自动控制模块控制气温和CO2浓度,对照气室气温与室外温度一致,根据室内外气温差异通过主控电脑控制大功率空调达到目标温度。控制系统通过气室内的 CO2传感器采集室内的空气并测定其 CO2浓度,并将此数据传输到主控电脑,按照控制程序控制各气室电磁阀的开闭,将各个气室的CO2浓度控制在目标浓度,室内安装环流风机,保证室内气温与CO2浓度均匀一致。系统还进行空气湿度和土壤湿度的监测(该系统由邯郸冀南新区盛炎电子科技有限公司设计)。试验共设 4个处理:CK(CO2浓度为 400 μmol∙mol-1,正常气温)、C0T1(CO2浓度为 400 μmol∙mol-1,正常气温+2 ℃)、C1T0(CO2浓度 600 μmol∙mol-1,正常气温)、C1T1(CO2浓度为 400 μmol∙mol-1,正常气温+2 ℃)。
谷子于2018年4月6日播种于直径28 cm高26 cm的桶中,每个桶随机撒播,播完之后覆土1 cm左右,桶下留有3个孔进行排水。每个处理4次重复。谷子出苗长出 3片真叶后间苗,每盆留苗 10株。及时浇水,防止干旱,浇水时间在早上和傍晚进行,保证无干旱胁迫。谷子从出苗开始一直到收获整个生长期全天24 h均进行CO2浓度及温度控制,各气室其他管理措施一致。
1.3 测定指标和方法
1.3.1 光合作用的测定
在谷子灌浆期(2018年5月27日),使用便携式光合气体分析仪(Li 6400,Li-CorInc,Lincoln NE,USA)进行了一次光合生理测定,包括净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr),并计算水分利用效率(WUE),WUE=Pn/Tr。使用内置红蓝光源,光量子通量密度(PPFD)为 1400 μmol∙m-2∙s-1(测定前随机取两株谷子进行光响应曲线测定,1400 μmol∙m-2∙s-1为测定日谷子饱和光强)。测定时叶室内CO2浓度设定同试验设计,正常气温气室内的叶温设定为25 ℃,升高气温气室内叶温设定为 27 ℃。对每个处理分别选取有代表性的植株 4株(每桶随机取 1株测定),取完全展开的旗叶,测定时间为09:00—12: 00(王晨光等,2015)。
1.3.2 形态指标及生物量的测定
在谷子完全成熟后,即2018年6月24日将各气室内的植株地上部分收获,风干后进行考种。对所有谷子植株进行株高、茎直径、穗长、节数、茎质量、叶质量、穗质量、粒质量和地上部分生物量等进行测量。脱粒后测定粒质量,并计算谷子产量及生物量。
1.4 统计分析
以 Excel 2010进行数据整理及图表绘制,以SPSS 20.0统计软件中的方差分析进行显著性检验。以SPSS 20.0进行二因素方差分析,采用一般线性模型,以CO2、温度为固定因子,以试验重复作为随机因子。
2 结果与分析
2.1 大气CO2浓度和气温升高对谷子净光合速率的影响
大气CO2浓度和气温升高对谷子净光合速率的影响如图1所示。与对照相比,大气CO2浓度升高(C1T0)使谷子的净光合速率显著增加了 19.09%,气温升高(C0T1)对谷子净光合速率没有显著影响。升温和升CO2复合处理(C1T1)条件下谷子净光合速率显著增加了16.95%。CO2对谷子净光合速率的影响达显著水平。升温和升CO2复合处理对谷子净光合速率交互作用不显著。
2.2 大气CO2浓度和气温升高对谷子气孔导度的影响
气温升高(C0T1)使谷子气孔导度显著增加145.33%;大气 CO2浓度升高(C1T0)使谷子的气孔导度下降16.80%;升温和CO2复合处理(C1T1)条件下谷子的气孔导度显著增大了条件下谷子的气孔导度显著增加73.62%。温度、CO2及其交互作用均达到显著水平(图2)。
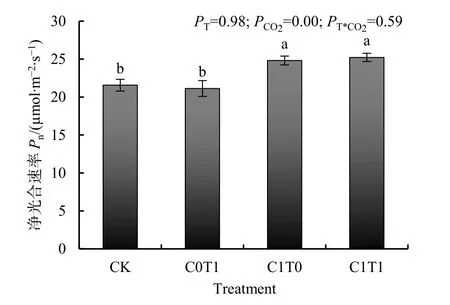
图1 大气CO2浓度和气温升高对谷子净光合速率的影响Fig.1 Effect of elevated atmospheric CO2 concentration and temperature on photosynthetic rate of millet
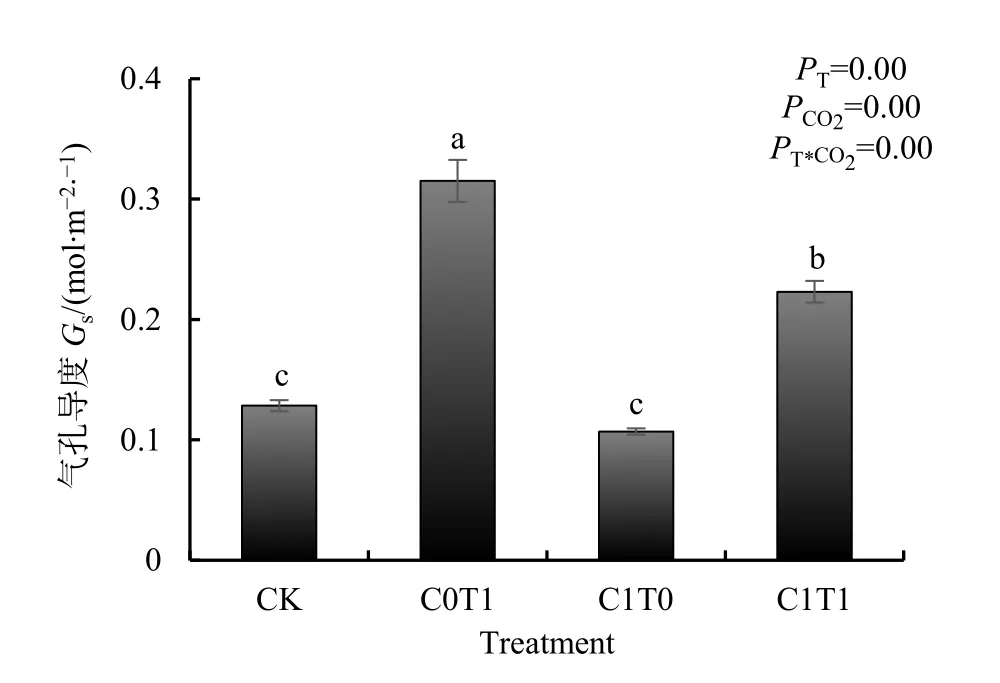
图2 大气CO2浓度和气温升高对谷子气孔导度的影响Fig.2 Effect of elevated atmospheric CO2 concentration and temperature on stomatal conductance of millet
2.3 大气CO2浓度和气温升高对谷子蒸腾速率的影响
气温升高(C0T1)使谷子蒸腾速率显著增加92.19%;大气CO2浓度升高(C1T0)使谷子蒸腾速率显著下降了 9.70%;升温和升 CO2复合处理(C1T1)条件下谷子的蒸腾速率显著增加 45.85%。温度、CO2及其交互作用均达到显著水平(图3)。
2.4 大气CO2浓度和气温升高对谷子水分利用率的影响

图3 大气CO2浓度和气温升高对谷子蒸腾速率的影响Fig.3 Effect of elevated atmospheric CO2 concentration and temperature on Trmmol of millet
气温升高(C0T1)使谷子水分利用率显著减少了49.32%;大气CO2浓度升高(C1T0)使谷子水分利用率显著增加了29.46%;升温和升CO2复合处理(C1T1)条件下谷子水分利用率减少了19.52%。温度、CO2对谷子水分利用率影响均达到显著水平。升温和升 CO2对水分利用率的交互作用不显著(图4)。

图4 大气CO2浓度和温度气温升高对谷子水分利用率的影响Fig.4 Effect of elevated atmospheric CO2 concentration and temperature on WUE of millet
2.5 大气CO2浓度和气温升高对谷子形态指标的影响
大气CO2浓度和气温升高对谷子形态指标的影响由表1所示。与对照相比,气温升高(C0T1)谷子株高、穗长、茎直径分别显著增加了 62.29%、63.51%和59.87%,节数无明显变化;与对照相比,大气CO2浓度升高(C1T0)谷子株高、穗长、茎直径和节数分别显著增加了35.18%、32.75%、25.80%和21.54%,;升温和升CO2复合处理(C1T1)条件下谷子的株高、穗长和茎直径分别增加了58.24%、42.69%和131.89%,茎数无明显变化。温度及两者交互作用均对上述形态指标均达到显著水平。CO2处理对谷子的株高及茎直径达显著水平。

表1 大气CO2浓度和气温升高对超早熟谷子形态指标的影响Table 1 Effect of elevated atmospheric CO2 and temperature concentration on growth of millet
2.6 大气CO2浓度和气温升高对超早熟谷子生物量和产量的影响
大气CO2浓度和气温升高对谷子生物量的影响如表2所示。与对照相比,气温升高(C0T1)谷子的叶质量、茎质量、穗质量和地上部分生物量分别增加73%、77%、70%和72.3%。大气CO2浓度升高(C1T0)谷子的叶质量、茎质量、穗质量和地上部分生物量分别增加了 21.5%、70.8%、19.4%和32.3%。升温和升 CO2复合处理(C1T1)条件下谷子的叶质量、茎质量、穗质量和地上部分生物量分别增加了34.8%、80%、77.6%和73.3%。气温处理对谷子生物量的上述参数均产生显著影响;CO2处理对谷子叶质量和茎质量的影响达显著水平;气温和CO2交互作用对谷子茎质量和地上部分生物量产生显著影响。
大气CO2浓度和气温升高对谷子产量的影响如图5所示。与对照相比,气温升高(C0T1)谷子的产量增加了76.6%;大气CO2浓度升高(C1T0)使谷子产量增加了28.7%,但未达显著水平;升温和升CO2复合处理(C1T1)条件下谷子产量显著增加了 92.6%。气温处理对谷子产量影响显著。CO2处理和两者交互作用均未达显著水平。
3 讨论与结论
3.1 讨论

图5 大气CO2浓度和温度气温升高对谷子产量的影响Fig.5 Effect of elevated atmospheric CO2 concentration and temperature on yield of millet
CO2是作物进行光合作用的重要原料。大气CO2浓度升高,由于 C3作物和C4作物的碳同化途径不同,C3作物光合作用的反应是C4作物的3倍(Ainsworth et al.,2005),而C4作物的光合速率仅提高4%(Kimball et al.,1997),没有C3作物影响大。虽然谷子是 C4作物,但本研究发现,CO2浓度升高使谷子的净光合作用升高19.09%,较前人研究结果高。表明部分 C4植物在目前大气 CO2浓度下没有达到饱和(Wand et al.,1999),在大气CO2浓度升高后净光合速率仍有较大提升空间。随CO2升高,植物光合作用最适温度也会升高 5—10 ℃(Drake,1997;Conroy et al.,1994)。有研究发现,达到最适温度之前,温度升高对作物的光合速率有促进作用。本研究发现,气温升高2 ℃对谷子净光合无显著影响。可能是因为是目前气温已接近谷子最适温度。CO2浓度增加可提高 CO2对Rubisco酶结合位点的竞争能力,从而提高羧化速度,同时通过抑制光呼吸提高净光合效率(张兆斌,2009)。但随着温度升高,CO2可溶性降低,Rubisco特异性减小,光呼吸消耗增大(杨金艳等,2003)。CO2浓度和温度升高的相互作用对光合作用有重要影响(余峥等,2006)。本研究发现CO2和温度同时升高,谷子的净光合速率显著升高,但二者交互作用不显著。可能由于在气温升高后谷子呼吸作用的加强而提高了对碳的需求(Griffin et al.,2002),促进光合作用的增加。

表2 大气CO2浓度和气温升高对谷子生物量的影响Table 2 Effect of elevated atmospheric CO2 concentration and temperature on yield of millet
大气CO2浓度的升高也会影响植物叶片的气孔运动,低浓度大气CO2会促进气孔张开,高浓度的CO2能够使气孔关闭(Bernacchi et al.,2005)。大气 CO2浓度升高后植物叶片的气孔导度平均下降22%(Ainsworth et al.,2007)。气孔关闭后植物蒸腾作用将减少20%—27%(Kimball et al.,1994;康绍忠等,1999)。由于蒸腾速率和净光合速率增加,植物水分利用效率将升高(曹生奎等,2009)。本研究发现CO2浓度升高后谷子叶片的气孔导度和蒸腾速率显著下降,水分利用率显著升高,这与前人之前的研究结果一致;升温处理下谷子气孔导度增大,蒸腾速率提高,这可能是叶片对高温的适应(廖轶等,2002;Radin,1992)。同时升温和升 CO2处理谷子气孔导度和蒸腾速率升高,但小于仅升高气温处理,表明高CO2可以减少叶片气孔导度,减少升温造成引起的水分损失;同时升温和升CO2处理的水分利用率小于仅升高CO2处理,CO2升高可提高谷子叶片水分利用效率。升温和CO2复合处理对气孔导度、蒸腾速率交互作用显著,升高CO2浓度可减少升温造成的水分损失,提高谷子叶片水分利用效率。
大气CO2浓度升高会促进植物叶面积生长,提高单位叶面积的净光合速率,并提高植物水分利用率,从而有利于植物生长及作物产量的提高(Curtis et al.,1998;Drake,1997)。本研究发现,CO2浓度升高条件下谷子株高、茎直径、穗长和节数显著增加,穗质量、粒质量、叶质量、茎质量也显著增加,这与前人的研究结果一致。气候变暖使农业生产的不稳定性增加,作物产量波动加大。多数学者认为温度升高将对作物产量起到负效应(Schiermeier,2001)。本研究发现,谷子在气温升高的条件下,株高、茎直径和穗长和节显著增加,穗质量、粒质量、叶质量、茎质量还有地上部分生物量也显著增加。这样的差异可能主要由于温度敏感性和植物生长发育的最佳温度存在着种间差别(曹素珍,2018)。曹素珍人工模拟增温的手段研究了植物出苗特性、生长、繁殖对增温的响应,结果表明植物的生长繁殖对增温的响应会在不同的物种间存在差异(Coleman et al.,1992)。原因可能是,(1)温度的升高加快了有机质在土壤中的分解速度,也加快了植物根系对土壤养分的吸收速率以及植物各项生理活动的速率(马丽等,2017)。(2)在温度和CO2浓度升高的共同作用下,植物体的营养需求增加导致根系生长的加强(Lilley et al.,2001)。(3)升高温度,使作物出苗时间提前,进而延长了植物生长季的长度,这同样有利于植物生物量的积累。关于大气CO2浓度和气温同时升高的综合作用对农作物产量影响方面的研究还不多,但从已有研究结果看,还是比较乐观的。如Costa et al.(2006)在OTC环境中分析高温、高CO2条件下水稻产量,结果表明,提高CO2收获时总生物量提高 23%—37%,可使穗粒数显著提高 24%。即使在高温条件下,水稻仍然对CO2的提高有积极的响应(Kim et al.,2011)。Usami et al.(2001)对 2 年生细叶青冈栎(Cyclobalanopsis glaucaOerst.)幼苗进行研究发现,全年温度升高3 ℃和5 ℃,相对生长速率增大,年生物量生产分别增大53%和47%。温度和 CO2浓度升高复合处理,年生物量生产增大 2倍。但也有试验证明,温度升高将会减弱 CO2升高对作物增产的效果(Wheeler et al.,1996;Lal et al.,1998;Ruiz-Vera et al.,2015)。在本试验中,升温和升CO2复合处理条件下谷子生物量和产量提高。
3.2 结论
(1)大气CO2浓度、温度升高及其复合处理均有利于超早熟谷子的光合作用及产量的提高。
(2)大气CO2浓度升高可以降低谷子叶片气孔导度,减少叶片蒸腾速率,提高谷子水分利用效率。
未来气候变化对山西晋中地区谷子的影响主要为正效应。但本研究是在控制气室内进行的盆栽试验,且仅为一个生长季试验。后续需要不同试验平台、不同地区、多年度试验对作物响应气候变化进行深入研究。
猜你喜欢
农业工程学报(2022年3期)2022-04-16
当代陕西(2021年21期)2022-01-19
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
农学学报(2019年1期)2019-02-21
延河(2018年6期)2018-06-11
新课程·下旬(2017年7期)2017-08-14
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2010年3期)2010-11-16
