
藜麦CqNHX基因家族鉴定及表达模式分析
2020-08-26 01:57:48贾冰晨吴筱林张东亮田晓芹陈世华郭善利
烟台大学学报(自然科学与工程版) 2020年3期
王 宇,贾冰晨,吴筱林,张东亮,褚 晶,田晓芹,陈世华,郭善利
(烟台大学生命科学学院,山东 烟台 264005)
NHX基因编码Na+(K+)/H+转运蛋白,属于阳离子/质子逆向转运体(Cation/proton antiporter,CPA),能够将细胞内的Na+(K+)逆向转运至细胞外或将Na+(K+)区隔化,保证细胞质内维持一个较低的Na+水平,调控细胞内的pH平衡[1-4].将拟南芥NHX转入番茄中可显著提高番茄对盐害的耐受性[4],NaCl、PEG及ABA胁迫下杜梨PbNHX1的表达水平显著上调[5],过量表达来自荷花的NnNHX1提高了烟草耐盐水平[6],过量表达来自霸王的ZxNHX显著提高了百脉根叶片和根部Na+、K+和Ca+的积累,并降低细胞渗透势,显著提高了百脉根抗旱和抗盐能力[7].这一系列研究结果表明,NHX在调节植物对应答盐胁迫过程中起到至关重要的作用.
藜麦(Chenopodiumquinoa.Wild)为苋科藜亚科藜属的一年生草本植物,原产于安第斯山脉,因为其种子营养丰富被现代营养学家誉为“营养黄金”[8],藜麦不仅种子营养价值丰富,而且该植物还具有一系列独特优异的生物学特性,例如耐盐碱、耐贫瘠、抗干旱等.随着该植物高质量参考基因组的发布[9],藜麦已经成为研究植物耐盐机制的重要植物材料之一,因此,利用生物信息学方法对藜麦CqNHX基因家族进行鉴定,全面分析其基因结构、染色体定位、系统发育关系及编码蛋白的理化性质等信息,并研究藜麦CqNHX基因在不同组织及非生物胁迫下的表达模式,进一步认识藜麦耐受盐胁迫的遗传分子机制,为耐盐藜麦和耐盐作物遗传改良和种质创新等方面提供一定的理论基础.
1 材料与方法
1.1 CqNHX基因家族成员的鉴定和理化性质分析
为了鉴定藜麦基因组中CqNHX基因家族,本研究使用含有NHX基因保守蛋白结构域的HMM模 型(PF00999)在藜麦基因组中进行搜索,E-value 限定为 1×10-5,并根据搜索结果构建藜麦特异的NHX的 HMM 模型,根据新的 HMM 模型对藜麦基因组进行搜索,搜索结果通过 PFAM 网站(http://pfam.xfam.org/)进行验证.藜麦基因组下载自藜麦基因组数据库(http://www.cbrc.kaust.edu.sa/chenopodiumdb/).
1.2 NHX基因的系统进化关系及跨膜结构预测
下载水稻和拟南芥NHX氨基酸序列,利用 MAFFT 在线网站(https://mafft.cbrc.jp/alignment/software/)进行联配,提取保守部分,然后将这些序列导入 MEGA7.0软件,选用最大似然法进行系统发育进化树的构建.所用数据分别由以下数据库获得:拟南芥(https://www.arabidopsis.org/),水稻(http://www.ricedata.cn/index.htm).使用鉴定出的藜麦NHX氨基酸序列导入在线网站(http://www.cbs.dtu.dk/services/TMHMM-2.0/)进行跨膜结构预测.
1.3 CqNHX基因结构及保守基序分析
从藜麦参考基因组的基因注释信息提取鉴定出的CqNHX基因结构信息,导入网站(http://gsds.cbi.pku.edu.cn/)的在线工具 Gene Structure Dispaly Server(GSDS)进行分析,绘制CqNHX的基因结构图.利用鉴定出的CqNHX基因氨基酸序列使用 MEME 在线网站(http://meme-suite.org/tools/meme)进行 motif 的查找和鉴定,保守结构域上限设置为 10 且允许保守结构域重复出现.
1.4 CqNHX基因表达模式分析
藜麦转录组数据来源于已经报道的藜麦转录组测序结果,SRA登录号为SRP226463和SRP116149,使用Kallisto[10]计算表达水平,使用Excel对得到的数据进行Log2转换,使用TBtools工具包[11]绘制热图,使用Excel绘制柱状图.
2 结果与分析
2.1 CqNHX基因家族的鉴定和理化性质分析
使用生物信息学方法从藜麦基因组中鉴定得到10个NHX基因家族成员,根据CqNHX基因与拟南芥AtNHX基因的亲缘关系将其命名为CqNHX1—CqNHX8,CqNHX基因的蛋白质长度,等电点和相对分子质量等生化属性如表1所示.其中蛋白长度分布在510~1211个氨基酸残基之间,等电点分布在5.38(CqNHX6b)到 9.18(CqNHX3b)之间.
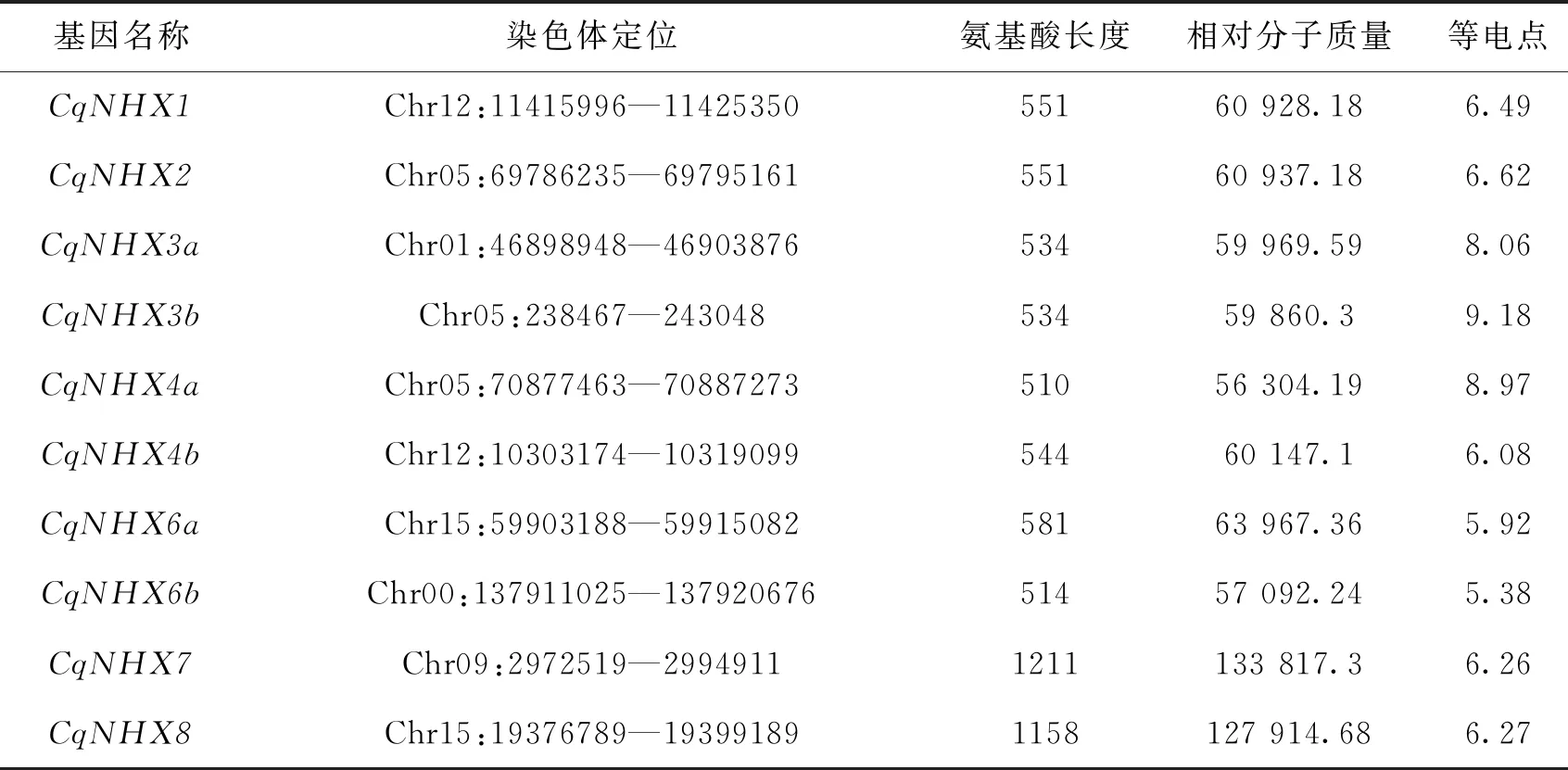
表1 CqNHX基因家族信息及理化性质
2.2 CqNHX基因的系统进化关系及跨膜结构预测
将拟南芥、水稻和藜麦的NHX氨基酸序列进行比对,使用MEGA7.0进行系统发育进化树的构建.结果表明:系统发育进化树将3个物种的NHX蛋白聚类为3个亚组(ClassⅠ、ClassⅡ和Class Ⅲ),ClassⅠ包含14个NHX基因,ClassⅡ包含2个藜麦CgNHX基因(CqNHX6a、CqNHX6b)、2个拟南芥AtNHX基因(AtNHX5、AtNHX6)以及1个水稻OsNHX基因(OsNHX5);ClassⅢ包含2个藜麦CqNHX基因(CqNHX7、CqNHX8)和2个拟南芥AcNHX基因(AtNHX7、AtNHX8)以及水稻OsSOS1基因(图1).
利用CBS在线网站对鉴定出的CqNHX蛋白进行跨膜结构预测.结果表明:CqNHX3a和CqNHX3b含有11个跨膜结构域,CqNHX4a、CqNHX4b和CqNHX7和CqNHX6b含有10个跨膜结构域,CqNHX6a仅含有9个跨膜结构域,而CqNHX1、CqNHX2、CqNHX7和CqNHX8中均含有12个跨膜结构域,对拟南芥AtNHX7(AtSOS1)的研究表明,N端密集的跨膜结构具有高度疏水性,而亲水性C端结构较长,残留在细胞质中,而CqNHX7和CqNHX8的跨膜结构域密集的集中在N端,且亲水C端结构较长,这与拟南芥AtNHX7的结构类似,表明CqNHX7/CqNHX8可能与AtSOS1具有类似的功能(图2).
2.3 CqNHX基因结构及其编码蛋白的保守基序
利用MEME 软件搜索并对NHX基因家族进行保守基序分析,保守基序上限设置为 10 且允许序列中同一基序重复出现,结果如图3,分析结果表明,CqNHX蛋白中保守基序长度较长,表明CqNHX成员间保守性较高,进一步对这些保守基序在NHX蛋白中的分布情况进行统计,发现系统发育进化树Class Ⅰ中的基因除CqNHX3a含有9个保守基序外,其余5个CqNHX基因的编码蛋白均含有10个保守基序,而Class Ⅱ和Class Ⅲ中的成员均含有5个保守基序.
对CqNHX基因家族成员进行基因结构分析,结果表明:除CqNHX3a和CqNHX3b不含有非编码区结构外,剩余8个CqNHX基因结构完整.对CqNHX基因家族成员进行外显子数量统计发现,CqNHX1、CqNHX2、CqNHX3a和CqNHX4b含有14个外显子结构,是所有CqNHX中外显子结构最少的,而CqNHX7中外显子数量最多,含有24个(图4).

表2 CqNHX蛋白的基序组成
2.4 CqNHX基因表达模式分析
RNAseq分析结果表明:ClassⅠ和ClassⅡ的6个CqNHX基因的表达具有明显的组织特异性,CqNHX1和CqNHX2除在种子外的所有组织中均检测到了表达,而CqNHX3a和CqNHX3b只特异性的在种子中表达(图5(a));CqNHX1和CqNHX2在藜麦地上部分组织中的表达受到干旱和高温的胁迫显著上调(图5(b)),而在根中这2个基因只受到干旱胁迫的诱导部分上调,对高温的响应不明显(图5(c));CqNHX3a和CqNHX3b在地上和地下部分组织中几乎检测不到,这与图5(a)展示的结果一致;CqNHX4a、CqNHX4b、CqNHX6a、CqNHX6b、CqNHX7和CqNHX8在藜麦地上部分组织中的表达受到干旱和高温胁迫显著下调,而在根中,CqNHX6a和CqNHX6b表达水平的变化不显著,CqNHX4a、CqNHX4b、CqNHX7和CqNHX8的表达水平只在干旱条件下有一定程度的上升,在高温胁迫下这些基因的表达水平几乎没有变化.
3 讨 论
随着研究的不断深入,大多数物种的NHX基因家族均被鉴定出来,例如玉米[12]、水稻[13]、大豆[14]、番茄[15]和小麦[16]中NHX基因家族均得到了鉴定和系统分析.本研究对藜麦、拟南芥和水稻NHX蛋白的系统发育进化关系分析表明,藜麦CqNHX7和CqNHX8与拟南芥AtNHX7(AtSOS1)和AtNHX8以及OsSOS1被聚到同一个分支,这表明CqNHX7、CqNHX8可能与AtSOS具有类似的功能并定位于细胞质膜上;系统进化分析中藜麦NHX基因与拟南芥NHX在进化距离上较水稻NHX更近,这可能与藜麦和拟南芥同属于双子叶植物,而水稻属于单子叶植物有关.对藜麦CqNHX蛋白的跨膜结构预测表明:CqNHX1、CqNHX2、CqNHX7和CqNHX8中均含有12个跨膜结构域,其中CqNHX7和CqNHX8蛋白在N端存在多次跨膜,这提示CqNHX7和CqNHX8可能在SOS信号途径[17]中起到重要作用.
植物在应对逆境胁迫的过程中有着多条途径,但在干旱、盐、高温等诱导的抗性方面具有相同或相似的基因作用机制[17].本研究对藜麦NHX基因的组织特异性和胁迫下的表达模式进行了探究,结果表明:CqNHX1、CqNHX2、CqNHX3a和CqNHX3b的表达具有明显的组织特异性,CqNHX3a和CqNHX3b特异的在种子中表达,因此CqNHX3a和CqNHX3b可能参与藜麦发育早期的K+利用和离子稳态过程中的生理过程.在干旱和高温的胁迫下,藜麦地上和地下部分NHX基因的表达水平具有较大差异,藜麦地上部分NHX基因对高温较为敏感,除CqNHX1和CqNHX2的表达受到干旱和高温胁迫显著上调外,其余NHX基因均受到高温和干旱的诱导而下调,这表明CqNHX1和CqNHX2在受到渗透胁迫后维持一个较高的表达水平,在Na+(K+)转运的过程中可能发挥重要作用.综上所述,本研究对藜麦NHX基因家族进行了系统的分析,为藜麦的分子育种提供了基因资源,同时也对NHX基因家族参与应答外界胁迫的分子机制研究奠定了一定的理论基础.
猜你喜欢
新医学(2023年10期)2023-12-09 15:04:51
南方医科大学学报(2022年3期)2022-04-13 01:51:26
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
福建质量管理(2019年21期)2019-11-28 09:01:44
建材发展导向(2019年11期)2019-08-24 06:35:26
心肺血管病杂志(2019年1期)2019-04-22 01:12:00
教学考试(高考生物)(2017年4期)2017-12-13 09:02:41
西南交通大学学报(2016年4期)2016-06-15 20:29:35
新疆农垦科技(2014年8期)2014-02-28 19:20:45
华东理工大学学报(自然科学版)(2014年2期)2014-02-27 13:48:43
