
circ-PTPN22在系统性红斑狼疮患者外周血单个核细胞中的翻译及意义
2020-08-18 10:00仲志婷苗青青蒋柱燕张梦洁唐隽
中华皮肤科杂志 2020年7期
仲志婷 苗青青 蒋柱燕 张梦洁 唐隽
1安徽医科大学附属解放军联勤保障部队第901 医院皮肤科,合肥230031;2陆军军医大学病理生理与高原病理学教研室,重庆400038
环状RNA(circRNA)是一种非编码RNA分子,没有5′端帽和3′端poly(A)尾部,通过共价键形成环状结构[1]。circRNA 可作为miRNA 的“海绵”来调节下游靶基因的表达[2],调节亲本基因转录[3],与RNA 结合蛋白相互作用[4],以及翻译蛋白质[5]。我们前期研究[6]采用高通量RNA测序,分析了3例系统性红斑狼疮(SLE)患者与3例健康对照外周血单个核细胞(PBMC)中circRNA 的差异表达,通过生物信息学分析筛选出4 个候选circRNA,并经实时荧光定量PCR(qRT-PCR)证实circ蛋白酪氨酸磷酸酶非受体22型(PTPN22)在SLE患者PBMC中表达降低,且与SLE 疾病活动指数(SLEDAI)评分呈负相关。本研究探讨circ-PTPN22的翻译能力及其翻译蛋白对Jurkat细胞活化和凋亡的作用。
对象与方法
一、研究对象
2019 年5 月10 日至9 月30 日分别于西南医院中西医结合风湿免疫科和体检中心收集6 例SLE女性患者(SLEDAI评分10 ~14分)和9例健康女性对照全血样本,年龄25 ~50 岁。本研究经解放军第105 医院(现更名为解放军联勤保障部队第901医院)伦理委员会批准( 伦理编号105LL20150308),所有受试者均签署知情同意书。
二、细胞来源及主要试剂
Jurkat 细胞系来源于美国ATCC 细胞库。RiboTM荧光原位杂交试剂盒(广州锐博生物技术有限公司),circ-PTPN22-cy3探针和18S-cy3对照探针均由广州锐博生物技术有限公司设计合成。内参甘油醛-3-磷酸脱氢酶(GAPDH)单抗(美国Bioss公司),山羊抗兔抗体、山羊抗小鼠抗体(北京中杉金桥生物技术有限公司),circ-PTPN22-FLAG 和circ-PTPN22-NC-FLAG 重组慢病毒由上海生博生物医药科技有限公司构建,circ-PTPN22-shRNAFLAG和circ-PTPN22-shRNA-NC-FLAG慢病毒由上海泰儿图生物科技有限公司合成。Dynabeads™Human T-Activator CD3/CD28 用于活化和扩增人T细胞(美国Thermo Fisher Scientific 公司)。藻红蛋白标记的抗人白细胞介素(IL)-2 抗体、7-AAD Viability Staining Solution(美国Biolegend 公司),小鼠抗FLAG®M2 单抗(美国Sigma-Aldrich 公司),ELISA 试剂盒(英国abcam 公司),RIPA 裂解液、一抗稀释液、二抗稀释液、Quick-Blocker 封闭液、2×十二烷基硫酸钠聚丙烯酰胺凝胶(SDS-PAGE)蛋白上样缓冲液(上海碧云天生物技术有限公司),ExpressPlus™PAGE Gels 4%~20%(南京金斯瑞生物科技有限公司),EasySep™人T细胞分离试剂盒、EasySep™人B 细胞分离试剂盒、EasySep™人NK 细胞分离试剂盒(加拿大STEMCELL Technologies 公司),eBioscience™Foxp3 转录因子染色液(美国Thermo Fisher Scientific公司)。
三、方法
1.PBMC分离:全血与磷酸盐缓冲液(PBS)及样本密度分离液体积比为1∶1∶1,805×g离心20 min,取白膜层,加PBS 10 ml,201×g 离心5 min 后弃上清液,得到PBMC。加入二甲基亚砜和胎牛血清(体积比1∶9),-80 ℃过夜,第2天转入液氮中保存。
2.荧光原位杂交法观察circ-PTPN22在健康人PBMC中的表达定位:针对circ-PTPN22剪接位点设计一段带有cy3 标记的探针circ-PTPN22-cy3,对照组探针为18S-cy3。从3 例健康人5 ml 全血样本中分离PBMC。50 μl PBS 重悬PBMC,取2 张多聚赖氨酸处理过的载玻片,分别标记为circ-PTPN22-cy3组和18S-cy3组。各取10 μl细胞悬液到载玻片,静置待干。然后依次用4%多聚甲醛室温固定10 min,预冷通透液4 ℃5 min,200 μl预杂交液37 ℃30 min,避光下将circ-PTPN22-cy3 和18S-cy3 两种探针各1.5 μl 分别加入100 μl 预热的杂交液混匀,弃去载玻片上的预杂交液,将探针混合物滴到含有细胞的载玻片上,放入湿盒,37 ℃避光过夜。第2天,42 ℃避光条件下,分别用杂交洗液Ⅰ、杂交洗液Ⅱ、杂交洗液Ⅲ洗涤,然后用4′,6-二脒基-2-苯基吲哚(DAPI)染色10 min,最后荧光显微镜下拍照。
3. Jurkat 细胞培养与慢病毒转染:用RPMI 1640培养基(含10%胎牛血清、1%青霉素链霉素)、在37 ℃5% CO2恒温孵箱中培养Jurkat 细胞,第2 天换液。为验证预测的circ-PTPN22 蛋白编码功能,分别将circ-PTPN22-FLAG、circ-PTPN22-NCFLAG、circ-PTPN22-FLAG + circ-PTPN22-shRNAFLAG、circ-PTPN22-shRNA-NC-FLAG 重组慢病毒[病毒载体含绿色荧光蛋白(GFP)基因]转入Jurkat细胞(circ-PTPN22-FLAG 组、circ-PTPN22-NCFLAG 组、circ-PTPN22-shRNA-FLAG 组、circ-PTPN22-shRNA-NC-FLAG 组),同时设置Jurkat 细胞对照组(仅正常培养)。采用12 孔板,每孔5 ×105个细胞,每组各3 个复孔,重组慢病毒感染复数(MOI)均为50。培养48 h后收集细胞,期间可根据细胞生长状态添加完全培养基和传代。
4.circ-PTPN22pro 抗体制备与鉴定:委托上海烈冰生物医药科技有限公司测定circ-PTPN22核酸序列。根据测序结果,委托杭州华安生物技术有限公司[动物试验许可证号:SYXK(浙)2017-0009]针对circ-PTPN22 剪接位点处的特异性氨基酸序列GNSNYLPVSLQELARP 合成肽段并制备多克隆抗体,然后使用亲和层析柱纯化抗体。采用常规多克隆抗体制备方法,选用2 只体重2.5 kg 左右的健康新西兰雄性白兔(编号为4137、4138),采用多点皮下注射各免疫3次,首次免疫后第14天进行2次免疫,第2 与第3 次免疫间隔7 d,第3 次免疫后第7 天,于兔耳中动脉采少量全血提取血清进行间接ELISA检测。检测合格后,进行加强免疫,7 d后可采集全血。采用转染了circ-PTPN22 的Jurkat 细胞鉴定所制备的多克隆抗体,将转染的Jurkat细胞裂解上清液经SDS-PAGE电泳后,用所制备兔抗血清作为一抗进行Western印迹法分析。
5. 间接ELISA 法检测制备的兔抗circ-PTPN22pro 抗体效价:按ELISA 试剂盒说明书依次进行包被抗原(合成的目的肽段)、1×TBST缓冲液洗涤、TBST配制的1%牛血清白蛋白(BSA)封闭,加入稀释的待检样品(即制备的抗体)37 ℃反应1 h、洗涤2 次。设置阳性对照孔(阳性血清)与阴性对照孔(BSA)。然后加辣根过氧化物酶标记二抗羊抗兔抗体孵育,1×TBST 缓冲液洗3次。加底物显色,2 mol/L硫酸终止反应后,于酶标仪450 nm波长处读取吸光度。各实验孔与BSA阴性对照孔A450比值>2.1时,为抗体滴度判定终点。
6.磁珠分选人PBMC:根据EasySep™Human T Cell Isolation Kit,分别用250 μl EasySep™缓冲液重悬3 例健康对照和3 例SLE 患者各15 ml 全血提取的PBMC,并用细胞计数板计数,每管加入50 μl/ml Isolation Cocktail,混匀后室温等待5 min,振荡30 s后每管加入40 μl/ml RapidSpheres™,用EasySep™缓冲液补齐至2.5 ml,混匀后放入配套的磁铁中,室温静置3 min,垂直将细胞悬液倒入另一新流式管中,将新管放入磁铁中,静置3 min,同样的方法倒出,得到分选后的T 细胞。参照上述方法分选B、NK细胞。用相应的抗体检测细胞纯度。
7. Western 印迹法检测细胞中circ-PTPN22pro蛋白表达:按照“3.Jurkat 细胞培养与慢病毒转染”中方法转染48 h 后,收集各组Jurkat 细胞,RIPA 裂解液提取总蛋白。加入2 × SDS 蛋白上样缓冲液(1∶1),煮沸变性。每孔上样量30 μg,160 V恒压电泳35 min,25 V 25 mA 6 min 半干转到聚偏二氟乙烯膜上,室温封闭30 min,分别加一抗小鼠抗FLAG标签抗体(1∶1 000)、兔抗人GAPDH抗体(1∶4 000)4 ℃孵育过夜。TBST 洗膜5 次,每次6 min;加二抗山羊抗兔、山羊抗鼠抗体(均为1∶4 000)室温摇晃孵育1 h,TBST 洗膜3 次,每次10 min,然后用Bio-Rad 成像仪曝光拍照。使用Image Lab 6.0 软件计算每组circ-PTPN22pro 条带与GAPDH 条带的调整灰度值之比。此外,采用上述方法检测3例健康对照和3 例SLE 患者PBMC 及另3 例健康对照和3 例SLE 患者T、B、NK 细胞亚群中circ-PTPN22pro 表达,一抗为制备的兔抗人circ-PTPN22pro 抗体(1∶500),二抗为山羊抗兔抗体。
8. 流式细胞仪检测circ-PTPN22 对细胞活化和凋亡的影响:按照“3.Jurkat 细胞培养与慢病毒转染”中方法将circ -PTPN22、circ-PTPN22-NC、circ-PTPN22 - shRNA、circ - PTPN22 -shRNA-NC 慢病毒分别转染Jurkat 细胞(circ-PTPN22 组、circ-PTPN22-NC 组、circ-PTPN22-shRNA 组、circ-PTPN22-shRNANC 组)24、48 和72 h 后,每孔加Dynabeads ™Human T-Activator CD3/CD28 4 μl,恒温孵育6 h 后收集转染的Jurkat 细胞。分别用200 μl PBS重悬,各组取1×106细胞,加入200 μl穿孔液室温20 min,500 μl 通透缓冲液453 × g 离心8 min,洗涤2次。重悬后每管加入1 μl藻红蛋白抗人IL-2抗体,室温避光20 min后洗涤2次,用含2%胎牛血清的PBS重悬细胞,流式细胞仪检测IL-2阳性细胞比例,以检测细胞活化状态。此外,收集各组转染48、72、96 h后的Jurkat 细胞,检测前10 min每管加7-AAD Viability Staining Solution 2 μl,流式细胞仪检测细胞凋亡率,即7-AAD阳性细胞比例。
9.统计学方法:应用SPSS25.0 软件,采用两因素重复测量方差分析分别分析过表达和干扰circ-PTPN22 不同时间对Jurkat 细胞活化及凋亡的影响。采用GraphPad Prism8 软件,两组计量资料的比较采用两独立样本t检验,P<0.05认为差异有统计学意义。
结 果
一、circ-PTPN22在健康人PBMC细胞中的定位
荧光原位杂交法显示,circ-PTPN22 和阳性对照18S 均主要表达在健康人PBMC 细胞质内,见图1。
二、circ-PTPN22的翻译能力预测
测序结果显示,circ-PTPN22基因全长1 429 bp,含9 个外显子。为预测该基因是否具有翻译蛋白的能力,通过circRNADb 网站(http://reprod.njmu.edu.cn/circrnadb)进行比对,预测出circ-PTPN22 第718 ~838位碱基共121 bp的内部核糖体进入位点(IRES)序列(图2A 绿色序列)。因为测得的circ-PTPN22全长为1 429 bp,所以排除1 399 ~1 507 bp的潜在IRES 序列。翻译能力预测见图2B,ORF 框从910 bp 跨剪接位点至50 bp 处,潜在翻译的蛋白质序列共189 个氨基酸,将其命名为circ-PTPN22pro;GNSNYLPVSLQELARP 序列为该编码蛋白不同于全长PTPN22 蛋白的特异氨基酸序列(图2B)。

图1 荧光原位杂交法检测circ-PTPN22在健康人外周血单个核细胞中的定位 图像融合后可见circ-PTPN22与18S均主要表达在细胞质中

图2 circ-PTPN22蛋白编码预测 2A:circ-PTPN22基因的预测编码序列;绿色部分代表IRES序列,灰色部分代表ORF框,跨剪接位点翻译,红色标记ATG、TGA分别为翻译的起始密码子和终止密码子;2B:circ-PTPN22pro的预测氨基酸序列,序列的红色部分是不同于全长PTPN-22蛋白的独特氨基酸序列
三、circ-PTPN22pro多克隆抗体制备及鉴定
ELISA 结果显示,3 次免疫后,2 只兔血清效价均达到1∶256 000。Western 印迹法鉴定circ-PTPN22pro 多克隆抗体显示,4 137 号兔血清的特异性较好,相对分子质量为20 000左右的目的条带显示清晰,但未检测到全长型PTPN22pro(相对分子质量98 000)(图3),提示circ-PTPN22pro 多克隆抗体制备成功,特异性针对circ-PTPN22pro。因此,在后续临床相关实验中使用4 137 号兔血清进行实验。
四、circ-PTPN22编码功能验证
转染重组慢病毒后48 h,circ-PTPN22-FLAG组、circ-PTPN22-NC-FLAG组、circ-PTPN22-shRNAFLAG 组、circ-PTPN22-shRNA-NC-FLAG 组细胞均有绿色荧光表达(图4A),说明重组慢病毒转染成功。Western 印迹结果显示,circ-PTPN22-FLAG 组circ-PTPN22pro 表达明显高于circ-PTPN22-NCFLAG 组和细胞对照组,且相对分子质量约为20 000;circ-PTPN22-NC-FLAG 组及细胞对照组有少量表达,提示Jurkat 细胞内源性表达circ-PTPN22pro(图4B)。与circ-PTPN22-FLAG组相比,circ-PTPN22-shRNA-FLAG 组circ-PTPN22pro 表 达明显降低,见图4C。
五、circ-PTPN22对细胞活化和凋亡的影响
转染24、48、72 h后,流式细胞仪检测显示,circ-PTPN22 组IL-2 表达(22.20% ± 8.92%,31.10% ±5.88%,53.20% ± 10.25%)均低于circ-PTPN22-NC组(30.90% ± 11.00%,51.23% ± 10.70%,69.67% ±9.00%);circ-PTPN22-shRNA组IL-2表达(35.57%±8.79%,78.10%± 10.08%,88.63%± 3.89%)均高于circ-PTPN22-shRNA-NC 组(26.73% ± 4.92%,41.03% ± 10.45%,41.33% ± 4.96%),见图5A、5B。两因素重复测量方差分析显示,circ-PTPN22 组与circ-PTPN22-NC 组间差异有统计学意义(F =284.881,P=0.003);circ-PTPN22-shRNA 组与circ-PTPN22-shRNA-NC 组间差异亦有统计学意义(F=293.818,P=0.003)。
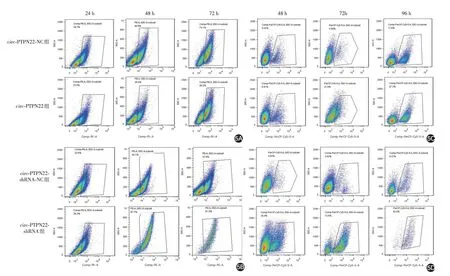
图5 流式细胞仪检测circ-PTPN22 对Jurkat 细胞活化和凋亡的影响 转染24、48 和72 h 后,circ-PTPN22 组IL-2 表达均明显低于circ-PTPN22-NC组(5A),circ-PTPN22-shRNA组IL-2表达均明显高于circ-PTPN22-shRNA-NC组(5B);转染48、72、96 h后,circ-PTPN22组细胞凋亡率均高于circ-PTPN22-NC组(5C),circ-PTPN22-shRNA组细胞凋亡率亦明显高于circ-PTPN22-shRNA-NC组(5D)
转染48、72、96 h后,流式细胞仪检测显示,circ-PTPN22 组细胞凋亡率(7.08% ± 1.33%,19.17% ±3.29%,30.03% ± 9.24%)高于circ-PTPN22-NC 组(5.01% ± 1.52% ,5.63% ± 0.81% ,14.22% ±3.96%),且circ-PTPN22-shRNA 组细胞凋亡率(24.63% ± 10.58%,59.17% ± 15.64%,84.87% ±2.91%)亦明显高于circ-PTPN22-shRNA-NC 组(6.59% ± 3.37% ,5.33% ± 1.65% ,15.42% ±7.31%),见图5C、5D。两因素重复测量方差分析显示,circ-PTPN22 组与circ-PTPN22-NC 组间差异有统计学意义(F = 81.287,P = 0.012);circ-PTPN22-shRNA 组与circ-PTPN22-shRNA-NC 组间差异亦有统计学意义(F=111.813,P=0.009)。
六、circ-PTPN22pro 在 健 康 对 照 和SLE 患 者PBMC中的差异表达
Western 印迹结果显示,SLE 组PBMC 中circ-PTPN22pro 表达低于健康对照组,几乎不表达(图6A)。进一步用磁珠分选后结果显示,SLE 组T、B细胞中circ-PTPN22pro 表达(0.398±0.147、0.072±0.027)均显著低于健康对照组(1.090 ± 0.365、0.512 ± 0.157;t 值分别为3.047、4.806;P 值分别为0.038、0.009),SLE组与健康对照组NK细胞中circ-PTPN22pro 表达差异无统计学意义(0.139 ± 0.147比0.088±0.032;t=0.582,P=0.592),见图6B。
讨 论
一些有IRES 序列及ORF 框的circRNA 具有编码蛋白能力[7]。我们在circRNADb数据库比对circ-PTPN22序列,发现circ-PTPN22有跨剪接位点翻译的ORF 框,并在Jurkat 细胞中检测出circ -PTPN22pro表达。通过荧光原位杂交法,我们检测出circ-PTPN22 主要在细胞质中表达,提示circ-PTPN22 可能主要在细胞质中发挥作用。与circ-PTPN22-NC 组相比,过表达circ-PTPN22 可抑制Jurkat 细胞活化,促进Jurkat 细胞凋亡;而干扰了circ-PTPN22 的表达可促进细胞活化,同时促进细胞凋亡。推测细胞凋亡的原因可能是干扰circ-PTPN22 表达后,circ-PTPN22pro 蛋白表达降低,导致细胞过度活化诱导了细胞凋亡[8]。

图6 系统性红斑狼疮(SLE)患者外周血单个核细胞(PBMC)及其T、B、NK 细胞亚群中circ-PTPN22 蛋白表达 6A:PBMC 中circ-PTPN22pro表达 3例SLE患者均明显低于3例健康对照;6B:T、B细胞中circ-PTPN22pro表达 SLE患者均明显低于健康对照,但两组NK细胞中circ-PTPN22pro表达无明显差异
通过制备兔抗circ-PTPN22pro 特异性多克隆抗体,我们检测了SLE患者和健康对照PBMC及其T、B、NK细胞亚群中circ-PTPN22pro的表达。结果显示,SLE 患者组PBMC 中circ-PTPN22pro 表达明显低于健康对照组,甚至不表达;SLE 患者组T、B细胞亚群中circ-PTPN22pro 表达亦显著低于健康对照组,而两组NK 细胞中circ-PTPN22pro 表达无明显差异。提示circ-PTPN22 和circ-PTPN22pro 蛋白可能对SLE的发生发展有一定作用,是影响SLE疾病的重要靶标分子之一。
PTPN22是一种淋巴特异性磷酸酶(Lyp),可抑制T细胞活化;当PTPN22第620位精氨酸(R)突变成色氨酸(W)时,PTPN22活性增强,可抑制T 细胞受体(TCR)的信号传导,从而促进自身免疫的发展[9]。PTPN22突变体PTPN22-R620W(LypW)在多种自身免疫疾病中起重要作用,比如SLE[10]、类风湿关节炎[11]等。SLE患者血清干扰素(IFN)-α水平升高,且IFN 驱动的下游基因表达增高[12]。然而,携带PTPN22 突变体LypW 的SLE 患者与非携带SLE患者血浆IFN-α水平无明显差异,但携带LypW的患者产生Toll 样受体(TLR)7 诱导的Ⅰ型IFN 能力降低,提示携带LypW 的SLE 患者可能存在Toll样受体或浆细胞样树突细胞依赖的宿主防御或抗炎功能缺陷[13]。PTPN22 是circ-PTPN22 的亲本基因,推测circ-PTPN22 可能与其亲本基因之间有相互作用或者具有类似PTPN22的作用。在后续研究中,我们将对circ-PTPN22与PTPN22相互作用进行深入研究,探索circ-PTPN22的作用机制,研究circ-PTPN22 或circ-PTPN22pro 蛋白在SLE 中的作用及机制。
志谢陆军军医大学免疫学教研室杨玓老师和田志强老师;西南医院中西医结合风湿免疫科和体检中心
利益冲突所有作者均声明不存在利益冲突
猜你喜欢
材料与冶金学报(2022年2期)2022-08-10
临床肺科杂志(2021年4期)2021-12-23
现代临床医学(2021年4期)2021-07-31
科学与财富(2021年33期)2021-05-10
中国科技纵横(2021年24期)2021-03-02
北京大学学报(医学版)(2020年6期)2020-12-14
中国现代医药杂志(2020年10期)2020-12-14
作文成功之路·小学版(2020年6期)2020-07-27
蚕桑通报(2020年1期)2020-07-10
作文成功之路·小学版(2020年5期)2020-06-11
