
菊粉内切酶在毕赤酵母中的表达及酶学性质测定
2020-07-31 07:46罗文浩普元倩徐春萍杨健康
井冈山大学学报(自然科学版) 2020年3期
王 淮,罗文浩,普元倩,徐春萍,杨健康
(1.大理大学基础医学院,云南,大理 671000; 2. 中山大学光华口腔医学院,广东,广州 510080)
1998 年Uhm 等人从无花果曲霉(Aspergillus ficuum)中分离到了一种新的菊粉内切酶基因,命名为inu2[1]。该基因编码了一个493 个氨基酸的蛋白,分子量约54 KD。N 端含有的一个23 个氨基酸的信号肽序列,使得该蛋白可以分泌到胞外。毕赤酵母(Pichia pastoris)作为高效的外源蛋白表达系统,到目前为止已经有上百种外源蛋白成功地在该系统中得到表达[2-3]。作为使用广泛的外源蛋白表达系统,毕赤酵母表达系统有比较明显的优点:在基因工程上操作起来比较简单、可以比较高的密度进行培养发酵,相比原核表达系统它可以对表达的重组蛋白翻译后进行加工修饰等[4-5]。但是毕赤酵母中并不是所有想要表达的外源基因都是高表达,部分基因表达低的原因之一是由于不同种属间对使用的密码子的偏好性不同及外源表达序列没有合理的(G+C)含量造成的。研究表明,对外源蛋白编码基因的密码子进行优化调整,大部分情况下都能提高外源蛋白的表达水平[6-7]。因此,我们下载了无花果曲霉菊粉内切酶基因的编码序列,做了密码子优化后对基因进行了全合成,然后在毕赤酵母GS115 中利用分泌载体pPIC9K 进行了分泌表达,在上清渡中获得了很高的酶活。同时,我们对酶的酶学性质进行了测定,并对菊粉内切酶酶解底物菊粉的最终产物进行了高效液相色谱(HPLC)分析。
1 材料与方法
1.1 基因
下载无花果曲霉菊粉内切酶基因(inu2)的序列(NCBI 编号AJ006951),编码序列长度为1,551 bp,编码了一个516 个氨基酸的蛋白。N 端含有一个23 个氨基酸的信号肽,使得该蛋白可以分泌到胞外。因为表达载体pPIC9K 带有自己的信号肽序列,所以需要去除基因自己的信号肽,去除信号肽序列后,基因编码序列长度为1,482 bp。
1.2 方法
1.2.1 密码子优化及评估
使用在线优化软件JCat(http://www.jcat.de)对菊粉内切酶基因进行优化[8],JCat 根据原核生物和真核生物的密码子使用偏好不同对待表达基因的序列进行一定的优化调整。我们选择根据酿酒酵母(Saccharomyces cerevisiae)的密码子使用偏好对无花果曲霉的菊粉内切酶基因进行密码子优化。
调整优化后的序列用BioEdit 软件将DNA 序列翻译为蛋白质序列并对序列进行比对,比较优化前后的蛋白质序列,要确认蛋白质序列没有因为密码子优化而改变。因为蛋白翻译的效率同RNA 的二级结构关系密切,RNA 的二级结构通过影响蛋白的翻译从而影响了基因的表达水平,所以根据RNA的二级结构可以对基因能否高表达进行实验前的预测分析[9]。本研究使用RNA 二级结构分析软件ViennaRNA Web Services ( http://rna.tbi.univie.ac.at ) 中的RNAfold Server,得到密码子优化前后基因序列的RNA 二级结构[10]。
1.2.2 全基因合成及载体构建
由上海捷瑞生物工程有限公司进行全基因合成,两头含有EcoRⅠ和NotⅠ酶切位点。全基因合成后的基因片段构建于pGH 质粒中,pGH 质粒为Amp 抗性。抽提质粒后,用限制性内切酶EcoRⅠ和NotⅠ对pGH 质粒以及即将连入的pPIC9K 质粒进行双酶切,回收1.5 kb inu2 基因片段,并与pPIC9K 质粒进行连接,构建pPIC9K-inu2 质粒,并测序保证序列正确。
1.2.3 毕赤酵母转化、多拷贝菌株筛选及摇瓶培养
用BglⅡ限制性内切酶酶切pPIC9K-inu2 质粒使载体线性化,并采用LiCl 法转化毕赤酵母GS115细胞。涂布于MD 培养基上,30 ℃培养3 d。从MD 平板上挑取生长良好的菌体少许,在含有不同抗生素G418 浓度(2、3、4 mg/mL)的YPD 培养基上划线培养3 d。在不同G418 浓度都生长较好的克隆即为高拷贝菌种。
将具有多个拷贝的克隆分别接种到BMGY 摇瓶培养基内,在30 ℃培养24 h。离心收集菌体,将收集的菌体转接到BMMY 摇瓶培养基中,用甲醇诱导表达3 d 后,收集摇瓶的上清液。
1.2.4 菊粉内切酶酶学性质研究
1.2.4.1 酶活测定
采用SDS-PAGE 分析目的蛋白[11]。将收集的上清液稀释10 倍,取50 μL 稀释后的上清液加入450 μL菊粉标准溶液中(2%菊粉溶解在NaAC-HAC 的缓冲液中,pH6.0),55 ℃反应10 s。用二硝基水杨酸法(DNS 法)测定菊粉内切酶将菊粉酶解后产生的还原糖[12-13]。100 ℃高温煮沸10 min,使上清液中的菊粉内切酶失去活性并作为对照。
酶活力定义为1 min 内产生1 μmol 还原糖所需的菊粉内切酶为1 个酶活力单位(U)。
1.2.4.2 酶的最适反应温度和热稳定性
分别在不同温度(40、45、50、55、60、65、70 ℃)测定酶活,研究菊粉内切酶的最适反应温度;研究该酶的热稳定性,将酶在不同温度(40、50、60 ℃)放置,在12、24、36、48、60、72 h 取样分别测定酶剩下的酶活,从而确定酶的热稳定性。
1.2.4.3 酶的最适反应pH 值
分别在不同的pH(4.0~9.0)条件下测定菊粉内切酶酶活,研究确定该酶的最适反应pH 值。
1.2.5 菊粉内切酶水解产物分析
用醋酸缓冲液配制 2%(w/v)菊粉溶液,在450 μL 底物溶液中加入50 μL 稀释10 倍的酶液,置于50 ℃中水解反应,反应12 h 后在沸水中加热,使酶活性丧失,停止反应。12,000 rpm 离心10 min后用超滤管超滤去除蛋白,取超滤到管中的液体用于HPLC 分析。在安捷伦液相色谱仪上分析,采用视差折光检测器检测,以乙腈和水为流动相,比例为70:30,流速为1 mL/min,柱温设为40 ℃,进样量10 μL[14]。用葡萄糖、果糖、蔗糖、蔗果三糖、四糖、五糖配制标准品溶液,各组分浓度均为1.0 mg/mL。
2 结果与分析
2.1 密码子优化结果
从NCBI 下载了inu2 的基因序列,用Jcat 在线软件优化后,原序列中的密码子都优化为最优密码子,同时优化时避免限制性内切酶酶切位点的产生。原始序列GC 含量为54%,密码子适应指数(CAI)为0.05,优化后序列GC 含量降低到52%,CAI 提高到0.73。RNA 二级结构会影响基因的蛋白表达水平,用软件RNAfold Server 预测优化前后的基因序列的二级结构见图1。从图可知,优化后的序列其结构分支减少了,可以推测优化后序列的RNA 二级结构较原来序列的结构更优,表达水平也可能更高。
2.2 菊粉内切酶酶活
将inu2 基因克隆到毕赤酵母表达载体pPIC9K上,测序保证序列正确,见图2。经BglⅡ线性化转化毕赤酵母GS115,抗生素G418 筛选高拷贝克隆。SDS-PAGE 分析表明,菊粉内切酶基因表达的蛋白大小正确(图3)。用DNS 法对菊粉内切酶的酶活进行测定,酶活力为420 U/mL。
2.3 酶学性质研究
将酶活的最大值设定为100%,在不同的温度下(40、45、50、55、60、65、70 ℃)测定酶活,作出酶的最适反应温度曲线,如图4 所示,酶的最适反应温度为55 ℃,在50~60 ℃范围内该酶活性较高;酶的热稳定性,酶在40 ℃条件下非常稳定,温浴72 h酶仍然可以保持60%以上的活性,而50 ℃下温浴12 h 后酶就仅剩下60%左右的活性,60 ℃下温浴12 h 后酶的活性基本丧失;在不同pH(4.0~9.0)条件下测定酶的活性,将最高酶活设定为100%,作出酶的最适反应pH 曲线,如图4 所示,该酶的最适反应pH 为6.0,在低于pH4.0 或高于9.0 时则几乎检测不到活性。

图2 密码子优化后序列测序图 Fig.2 Sequencing diagram of codon optimized sequence

图3 菊粉内切酶蛋白的SDS-PAGE 图谱 Fig.3 SDS-PAGE of endoinulinase protein
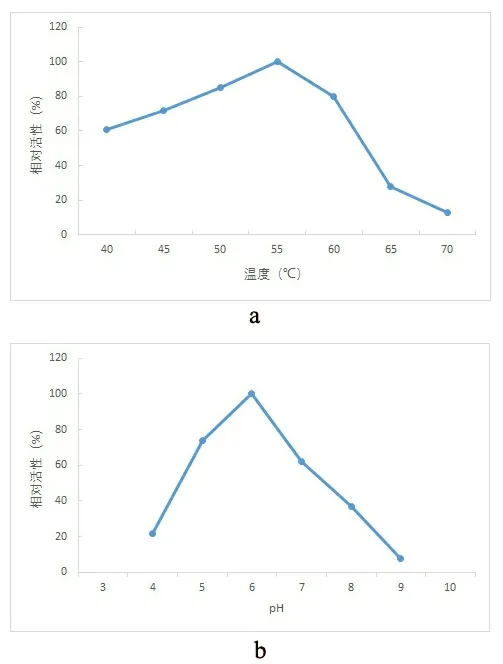
图4 菊粉内切酶的最适反应温度(a)和最适反应pH 值(b) Fig. 4 Optimal reaction temperature of endoinulinase (a) and optimum pH (b)
2.4 酶水解菊粉产物分析
酶水解菊粉产物的HPLC 图谱如图5 所示。由于只购买到聚合度为3-5 的低聚果糖,因此在缺乏标准品的情况下没有对聚合度大于5 的糖进行分析。从图5 可以看出酶水解菊粉的产物主要是聚合度为3-5 的低聚果糖,此外还有少量聚合度大于5的低聚果糖。

图5 菊粉水解产物HPLC 图谱 Fig.5 HPLC chromatogram of inulin hydrolysate
3 讨论
本研究利用异源基因表达技术对来自曲霉的基因在毕赤酵母中进行了分泌表达,并对菊粉内切酶基因inu2 密码子进行了优化,构建了毕赤酵母pPIC9K-inu2/GS115 菌株,其表达的酶活力为420 U/mL。还对inu2 基因在酿酒酵母中的表达进行过研究,但蛋白表达量和酶的活性都比较低,无法应用于生产[15]。这说明菊粉内切酶基因的表达受表达体系的影响较大,在不同体系中表达水平差异很大。外源基因表达效率受到密码子偏好性的影响,不同物种密码子偏好性不同,无花果曲霉与酵母就存在很大的差异,这种偏好性实质反应的是物种间不同转运tRNA 含量的差异。如果基因含有稀有密码子,而相应的tRNA 含量不足,就会导致基因表达水平低。同时,mRNA 的二级结构也会影响与核糖体的结合,密码子优化时也要考虑二级结构的影响。综合考虑基因密码子偏好性以及mRNA 二级结构,我们对无花果曲霉的菊粉内切酶基因进行了密码子优化,经实验证明表达量有了很大的提高。
毕赤酵母的表达载体pPIC9K 属于分泌型载体,表达的蛋白分泌到胞外,有利于酶的分离和纯化。该载体成功地表达了多种外源蛋白质,本研究也发现利用毕赤酵母以及表达载体pPIC9K 能够实现inu2 基因的高表达。inu2 基因有自身的分泌型信号肽序列,长度为23 个氨基酸,而表达载体pPIC9K本来就有α-factor 分泌信号肽。曾有研究报道[16],使用自身信号肽的菊粉内切酶在酶活上低于使用α-factor 分泌信号肽的菊粉内切酶。因此在本研究中通过去除inu2 基因自身23 个氨基酸残基的信号肽序列,使用表达载体的α-factor 分泌信号肽。
目前知道可以产生菊粉内切酶的微生物有很多,包括细菌、真菌、酵母菌和放线菌等。为了获得高酶活的菊粉内切酶,研究者从上述微生物中都克隆了菊粉内切酶基因并放入毕赤酵母中进行了表达,获得了高产的重组菌株。Wang 等人将从Aspergillus niger 9891 获得的菊粉内切酶基因克隆表达在毕赤酵母GS115 中,对转化子进行7 升高密度发酵,菊粉酶活力达到291 U/mL,酶活力是原始菌株的273 倍[17]。林胜强等对表达菊粉内切酶的毕赤酵母工程菌inu2-26 进行高密度培养条件优化,酶活最高可达570 U/mL[18]。我们构建的pPIC9K- inu2/GS115基因工程菌表达的菊粉内切酶酶活力为420 U/mL,并不是最高的。我们目前使用的是摇瓶发酵,如果下一步上发酵罐进行高密度补料诱导发酵,酶活还有进一步的提高空间,曾有文献报道发酵罐高密度发酵可以使酶活再提高2~3 倍[18-19]。
低聚果糖主要指蔗果三糖、四糖、五糖,一些研究将聚合度更高的蔗果六糖也称为低聚果糖。低聚果糖不会被人体消化道内的各种消化酶消化分解,所以可以作用于肠道双歧杆菌等有益菌,促进有益菌的生长,改善肠道的微环境,从而间接促进人体的健康。此外,研究发现低聚果糖可以改善人体脂质代谢,能够用于减肥和糖尿病的防治[20]。利用基因工程菌生产的菊粉内切酶酶解菊粉制备低聚果糖,这一方法仅仅需要进行一次酶解就可以生成纯度比较高的低聚果糖,生产成本比较低,因此该方法在低聚果糖生产上应该有比较好的前景。
猜你喜欢
食品与发酵工业(2022年6期)2022-04-01
食品与发酵工业(2022年3期)2022-02-23
河南工业大学学报(自然科学版)(2021年5期)2021-11-15
生物技术通报(2021年6期)2021-08-11
微生物学杂志(2020年5期)2020-12-23
浙江化工(2019年4期)2019-05-13
大众健康(2018年11期)2018-11-28
百科知识(2018年22期)2018-11-27
浙江农业学报(2017年3期)2017-04-08
饮食科学(2016年9期)2016-11-18
