
谷子12 种黄酮类代谢物合成通路分析
2020-07-09 02:57:12张丽玲郄倩茹罗韶凡牛文康朱喆标高雨柔李旭凯韩渊怀
山西农业大学学报(自然科学版) 2020年4期
张丽玲 ,郄倩茹 ,罗韶凡 ,牛文康 ,朱喆标 ,高雨柔 ,李旭凯 *,韩渊怀 *
(1.山西农业大学农学院,山西太谷030801;2.山西农业大学生命科学学院,山西太谷030801)
谷子属于禾本科狗尾草属,去皮后俗称小米。谷子起源于我国,种植历史悠久,是重要的杂粮作物之一[1]。谷子具有抗旱、耐瘠、耐贮藏的优点,主要种植在我国内蒙古、山西、河北等地[2,3]。谷子含丰富的蛋白质、维生素B2、钙、铁等物质,营养价值极高[4,5]。《本草纲目》多处记载:“粟米(小米)味咸淡,气寒下渗,肾之谷也之,虚热消渴泄痢,皆肾病也。渗利小便,所以泄肾邪也,故脾胃之病宜食之”;“养肾气,去脾胃中热,益气”,说明谷子具有药食同源的功能。黄酮代谢物属于植物次生代谢物中相对重要的一类代谢产物,其合成是通过查尔酮合成酶催化丙二酰辅酶A(malonyl-CoA)与4-香豆酰辅酶A,而后将基本的碳架结构提供给三叶豆甙、花翠素3-O-葡萄糖苷、异槲皮甙、山奈酚 3-O-芸香糖苷、野漆树苷、黄腐醇等物质。黄酮代谢物普遍存在于植物膳食中,尤以蔷薇科(苹果,梨)、芸香科(橙、柚、柑、橘)、禾本科(麦、稻、玉米、高粱)等植物中含量较为丰富,具有抗氧化、抗炎、降血糖、清除自由基等多种生理活性[6,7]。茶叶中黄酮类物质对心血管起保护作用,能够降低心血管疾病的发病率[8]。有研究表明,蒲公英的抗氧化性与其花中所含的黄酮类物质有关[9]。郑楠楠等[10]对10 个品种谷子营养及功能活性成分进行分析发现,张杂谷5 号中黄酮含量显著高于晋谷21等其他品种,除此研究外,目前对晋谷21 的黄酮类次生代谢物的研究较少,分析其代谢通路对发掘谷子功能性黄酮类物质及相关基因具有重要意义。
1 材料和方法
1.1 试验材料
晋谷21(山西名优品种,JG21)、牛毛白(白色籽粒农家种,NMB)、xiaomi(晋谷21 通过EMS 诱变后获得的超早熟材料)的籽粒及叶片等组织材料。
1.2 谷子转录组测序数据的处理
谷子的转录组数据来源于本实验室RNA-seq测序,将超早熟xiaomi(待发表)、牛毛白及晋谷21于2019 年播种于山西农业大学杂粮分子育种团队基地(山西省太谷县,北纬 37°25′13″,东经 112°35′26″)。超早熟 xiaomi,分别取萌发后 2 周的叶、出苗30 d 和80 d 的倒2 叶,授粉期的谷穗和抽穗2天后的谷穗,灌浆期的根、茎、旗叶及穗,以及培养3 d 后萌芽的种子;晋谷21 及牛毛白品种均只取S2、S4 两个灌浆时期未成熟的籽粒。取样后将所有样品立即冷冻在液氮中,随后根据制造商的说明采用RNAprep 纯植物试剂盒(天根生化科技有限公司)分离总RNA,送诺禾致源公司进行转录组测序。所有的测序原始数据fastq 序列文件利用FastQC[11]对其进行质控。数据过滤采用Trimmomatic[12]软件,得到 clean data。利用 HISAT2[13]软件将 clean data 比对到谷子基因组,SAMtools[14]进行 sam/bam 文件处理,featureCounts[15]获得数据的reads 计数,计算得到基因表达的TPM 值(transcripts per million)。
1.3 黄酮类代谢物的提取与测定
将谷子成熟后的籽粒冷冻干燥后利用高通量组织研磨器在30 Hz 条件下研磨1.5 min,称取100 mg 研磨好的粉末于70%的甲醇溶液中提取,之后将其放入4 ℃的冰箱中过夜8 h 以上,期间震荡3 次,以提高提取率。将样品离心(转速10 000 r·min-1,10 min),吸 取 上 清 液 ,再 用 微 孔 滤 膜(0.22 μm pore size)过滤样品,吸取 150 μL 于进样瓶中用于LC-MS/MS 分析。
HPLC 条件和参数:用于 LC-ESI-MS/MS 分析的仪器系统主要包括HPLC(Shim-pack UFLC SHIMADZU CBM20A system,http://www. shimadzu. com. cn)和 MS(Applied Biosystems 4000 Q TRAP,http://www. appliedbiosystems. com.cn/),分析条件如下:shim-pack VP-ODS C18 色谱柱(pore size 5.0 μm,length 2×150 mm),0.1%乙酸的超纯水水相),0.1%乙酸的乙腈有机相;洗脱梯度为:0 min 为 100∶0 V·V-1,20.0 min 为 5∶95V·V-1,22.0 min 为 5∶95 V·V-1,22.1 min 为 95∶5 V ·V-1,28.0 min 为 95∶5 V·V-1;流 速 为 0.25 mL·min-1;柱温为 40 ℃;进样量为 5 μL。分离的样品接着进入ESI-QTRAP-MS,进行质谱分析。
ESI-QTRAP-MS/MS 条件和参数:ESI 源的电喷雾离子源温度为500 ℃,IS 为5 500V,GSⅠ、GSⅡ以及 CUR 分别为 55 psi、60 psi 和 25 psi,CAD 设置为高。
1.4 谷子黄酮类代谢物的分析
根据晋谷21 及牛毛白灌浆后期籽粒的黄酮代谢组分析结果,结合已有代谢物信息,找到具有对人体有益的黄酮类物质及其代谢通路上的酶基因编号,将酶基因编号提交于至酶数据库https://www. enzyme-database. org/[16],找出 其 在 其 他植物中的基因序列,之后用该基因序列在phytozome[17]数据库 Blast 谷子蛋白库,找到谷子代谢通路酶基因。将晋谷21 及牛毛白籽粒灌浆后期所测的代谢物含量及其酶基因表达量用Tbtools[18]进行热图分析,绘制代谢通路图。
2 结果与分析
2.1 谷子黄酮类物质代谢通路
从图1 可见,代谢通路图中有10 个关键基因,除异樱花亭,其余5 个黄酮代谢物在晋谷21 中的含量要高于牛毛白。柚皮苷查尔酮是代谢通路中的第一个生成物,促使晋谷21 柚皮苷查尔酮含量高于牛毛白的关键基因有Seita. 9G142900、Seita. 8G139300;柚皮苷查尔酮通过E5.5.1.6 酶作用下生成柚皮苷,该过程中促使晋谷21 灌浆后期籽粒柚皮苷含量高于牛毛白的关键基因有Seita. 8G013300。之后柚皮苷分别在E2.1.1.231酶和E1.14.20.5 的作用下生成异樱花亭和芹菜素,其中促使牛毛白灌浆后期籽粒异樱花亭含量高于晋谷21、晋谷21 灌浆后期籽粒芹菜素含量高于牛毛白的关键基因分别是Seita. 1G045400;Seita. 3G026900、Seita. 9G342600、Seita. 9G561700;而后芹菜素经由CYP75A 酶生成木犀草素,在此过程中使晋谷21 灌浆后期籽粒木犀草素含量高于牛毛白的关键基因有Seita. 9G393400;木犀草素在O-methyltransferase 酶的作用下生成金圣草(黄)素,促使晋谷21 灌浆后期籽粒金圣草(黄)素含量高于牛毛白的关键基因有Seita. 7G073700、Seita.9G374200。除此之外,从代谢通路图中还可以看出:优质品种晋谷21 中多数基因的表达量显著高于品质较差的牛毛白,具有抗氧化、抗炎、调节脂肪等作用的柚皮苷查尔酮[19]和具降血脂、镇静、抗氧化、抗肿瘤的柚皮苷[20]是 6 个黄酮代谢物中含量较高的2 个物质。

图1 相关物质代谢通路图Fig.1 The pathway of related substance
2.2 三叶豆甙的生成
三叶豆甙为黄酮醇类别的一种物质,由山奈酚通过E2.4.1.234 酶生成。从图2 可以看出,晋谷21 的灌浆后期籽粒中该物质的含量明显高于牛毛白,从基因表达热图中可以看出各组织中低表达或不表达的基因较多,如Seita. 1G124800、Seita. 7G318900、Seita. 3G189600、Seita. 1G124900、Seita. 6G104300、Seita. 9G389900等 基 因 。Sei-ta. 1G125200除在旗叶、出苗 30 d 后的倒 2 叶与根中表达较高外,其余组织中表达都低;Seita. 1G041600基因在授粉期及灌浆期穗与旗叶中表达较高,其余组织中表达均低且相差无几;Seita. 5G127500、Seita. 1G057100、Seita. 9G086800、Seita. 5G308400、Seita. 4G082500等基 因 表 达较高,其中Seita. 5G127500表达具有组织特异性,除在根中不表达外,在晋谷21 与牛毛白灌浆期籽粒中表达也较低,在其余组织中表达均较高;Seita. 1G057100在晋谷21 与牛毛白籽粒中表达显著高于其他基因在籽粒中的表达,且在晋谷21 中表达量明显高于牛毛白。此外,Seita. 2G275500、Seita. 2G419800、Seita. 9G086800、Seita. 4G228200等4 个基因在晋谷21 灌浆后期籽粒的表达略高于牛毛白;Seita. 1G057100在晋谷21 中籽粒表达量明显高于牛毛白,该基因可能是决定晋谷21 灌浆后期籽粒中三叶豆甙含量明显高于牛毛白的主效基因。

图2 三叶豆甙的代谢图与热图Fig.2 Metabolism and heatmap of trifolin
2.3 花翠素3-O-葡萄糖苷的生成
花翠素3-O-葡萄糖苷是由花翠素通过BZ1 酶生成的,由图3 可以看出,其代谢物含量在晋谷21籽粒中略高于牛毛白,Seita. 8G192500、Seita.8G192600在各组织中表达均比较低;Seita. 2G324300、Seita. 4G043100、Seita. 8G117800、Sei-ta. 8G117700、Seita. 7G079000、Seita. 8G203200等在晋谷21 及牛毛白S2、S4 籽粒中表达较高,其中Seita. 8G203200与Seita. 8G117700具 有 组 织 特 异性,仅在根、灌浆期的茎秆中表达最高;Seita. 7G079000、Seita. 8G117800、Seita. 2G324300在晋谷21 灌浆后期的籽粒中表达高于牛毛白,其中Seita. 2G324300在晋谷21 灌浆后期的籽粒中表达显著高于牛毛白,说明Seita. 2G324300可能是晋谷21 籽粒中代谢物含量高于牛毛白的主效基因。

图3 花翠素3-O-葡萄糖苷的代谢图和热图Fig.3 Metabolism and heatmap of mirtillin
2.4 异槲皮甙的生成
异槲皮甙是由槲皮素通过E2.4.1.91 酶生成的物质。由图4 可以看出,该代谢物在晋谷21 与牛毛白的籽粒中含量相差无几。在该通路中,Seita. 9G269600、Seita. 5G308200、Seita. 5G308900、Seita. 2G351100、Seita. 8G154000、Seita. 2G276000等基因在各组织中表达均低;Seita. 2G324200是所有基因中表达最高的;Seita. 9G409500、Seita. 5G308500在根中表达具有特异性;Seita. 2G092900在灌浆期茎秆中表达具有特异性;Seita. 9G239700、Seita. 1G325500、Seita. 1G041500在灌浆期旗叶中表达具有特异性;Seita.5G309200在萌发2 周的叶及出苗后30 d 的倒2 叶中表达具有特异性;Seita. 2G324600在穗与旗叶中表达较高,其余组织中较低;Seita. 5G309300在萌发3 d的萌芽种子、灌浆期的穗与根中表达较高。该通路中大多数基因在晋谷21、牛毛白中表达相差不大,这可能导致了异槲皮甙在晋谷21 与牛毛白籽粒中含量相差无几。
2.5 黄腐醇的生成
黄腐醇是由去甲黄腐醇通过OMT1 酶生成的一种物质。由图5 可以看出,牛毛白的黄腐醇含量明显高于晋谷21。Seita. 8G108400、Seita. 6G088200、Seita. 8G110100在根中表达具有特异性;Seita. 6G055900在各组织中表达最高,且其在抽穗后2 d及授粉期的穗中表达最高;Seita.8G203300在发芽3 d 的种子中具有特异性;Seita. 1G372500、Seita. 7G022900、Seita. 8G210100表达较高,其中Seita. 8G210100在牛毛白灌浆后期的籽粒中表达高于晋谷21,可能是牛毛白中黄腐醇含量显著高于晋谷21 的主效基因。
2.6 芹菜素7-O-新橘皮糖苷的生成
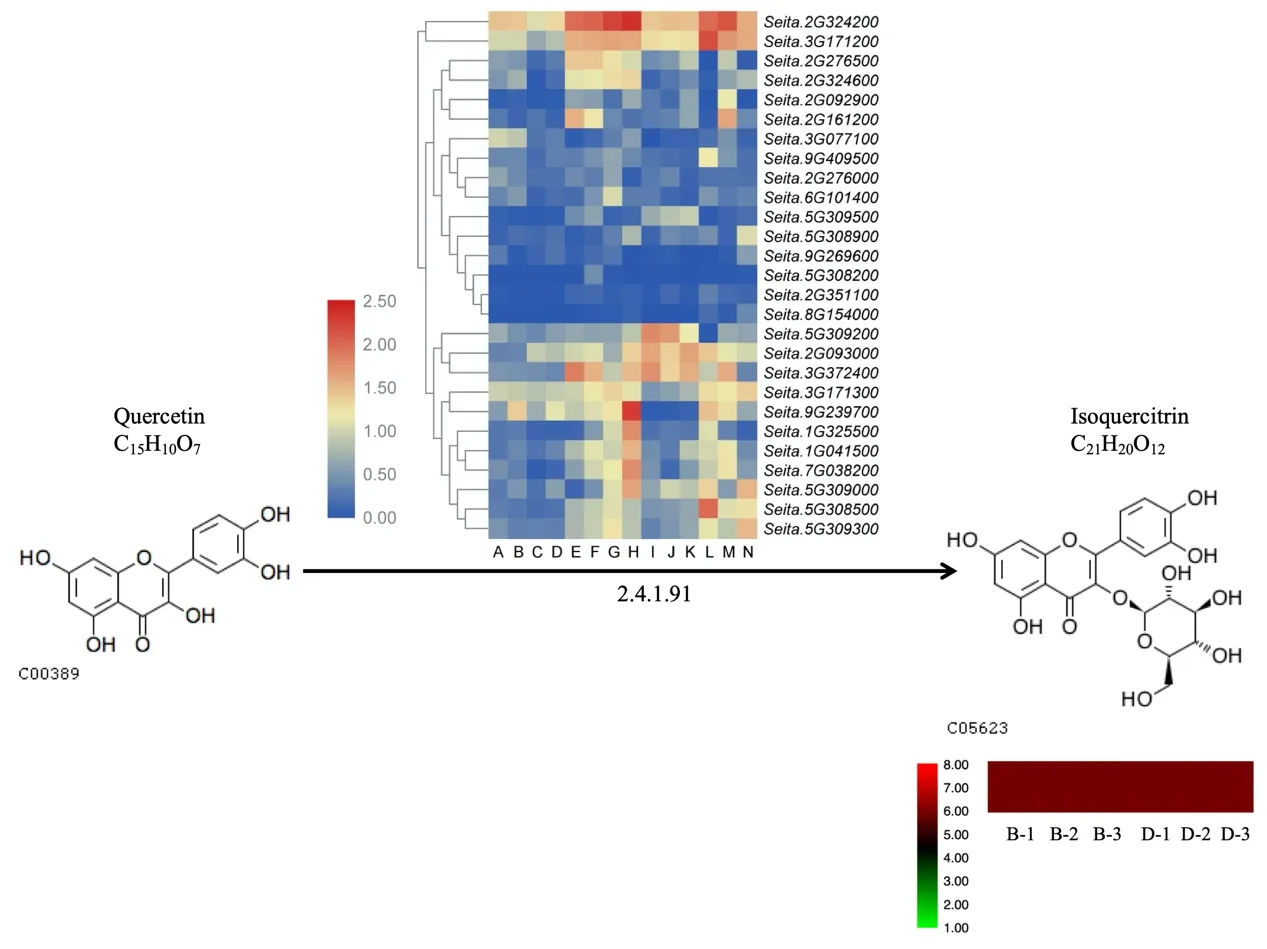
图4 异槲皮甙的代谢图和热图Fig.4 Metabolism and heatmap of isotrifoliin
芹菜素7-O-新橘皮糖苷又称野漆树苷,是由芹菜素-7-O-葡糖苷通过C12RT1 酶生成的一种物质,由图6 可以看出,牛毛白中该代谢物含量高于晋 谷 21。Seita. 1G041700、Seita. 2G324400、Seita. 8G134800、Seita. 5G124200、Seita. 4G045200、Seita. 7G194300、Seita. 2G324500、Seita. 3G287700、Seita. 7G264500、Seita. 7G171900等多个基因在除籽粒外的其余组织中表达较高;Seita. 7G031700、Seita. 1G304200、Seita. 2G192900、Seita. 7G139300、Seita. 6G191600、Seita. 7G171800等基因在各组织表达均最 低 ;Seita. 8G196100、Seita. 2G076300、Seita. 2G050100在根中表达较高;Seita. 2G179100在灌浆期的茎秆中表达具有特异性;Seita. 7G159300在叶、倒2 叶中不表达,在其余组织中表达较高;Seita. 2G076100在叶中表达最低;Seita. 9G493900在抽穗后2 d 的穗中表达较高;Seita. 4G127300在抽穗后2 d 及授粉期的穗及茎秆中表达具有组织特异性;Seita. 7G194900在萌发3 d 的萌芽种子中表达具有特异性;Seita. 2G427700、Seita. 9G542000、Seita. 7G194700在牛毛白灌浆后期籽粒的表达量比晋谷21 高,其中Seita.9G542000最显著,说明这可能是牛毛白中该代谢物含量高于晋谷21 的主效基因。
2.7 山奈酚3-O-芸香糖苷的生成
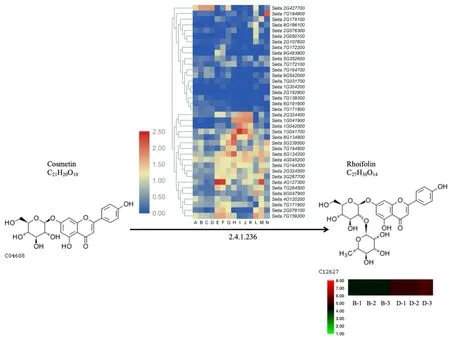
图6 野漆树苷的代谢图和热图Fig.6 Metabolism and heatmap of rhoifolin
山奈酚3-O-芸香糖苷是由紫云英苷通过FG2酶来生成的。由图7 可以看出,该物质在牛毛白中含量显著高于晋谷21。从图中可以看出,Seita. 2G222400、Seita. 3G189800、Seita. 3G189900、Seita. 4G177600、Seita. 5G392600、Seita. 8G192600、Seita. 9G095800等基因在组织中表达最低。Seita. 6G165400、Seita. 2G102600、Seita. 8G203200在根中表达具有组织特异性;Seita. 2G076000在根与茎秆中表达较高;Seita. 9G163100在2 周的叶及倒2 叶中表达具有特异性;Seita. 5G124300在旗叶中具有组织特异性;Seita. 3G190000在出苗30 d的倒 2 叶中表达较高;Seita. 9G125900除在 2 周叶及出苗30 d 的倒2 叶中表达较高外,在牛毛白灌浆后期籽粒中表达高于晋谷21,推测Seita. 9G125900可能是该物质在牛毛白中含量显著高于晋谷21 的主效基因。
3 讨论与结论
黄酮类物质对于缓解人类疾病具有明显作用,研究谷子中的黄酮类物质至关重要。本研究对谷子14 份籽粒及叶片等组织材料的转录组进行了分析,对晋谷21 及牛毛白灌浆后期籽粒黄酮代谢物及酶基因热图进行了分析,结果表明在黄酮代谢通路中有15个关键基因,分别为Seita. 1G057100、Seita. 2G324300、Seita. 9G125900、Seita. 9G542000、Seita. 8G210100、Seita. 9G142900、Seita. 8G139300、Seita. 8G013300、Seita. 1G045400、Seita. 3G026900、Seita. 9G342600、Seita. 9G561700、Seita. 9G393400、Seita. 7G073700、Seita. 9G374200,其中有 9 个关键基因促使7 个黄酮类物质在晋谷21 中含量高于牛毛白,有6 个关键基因促使4 个黄酮类物质在牛毛白中含量高于晋谷21。黄酮代谢物中柚皮苷查尔酮、柚皮苷是含量最高的黄酮代谢物,其中柚皮苷查尔酮具有抗氧化、抗炎、调节脂肪和性激素代谢的作用,柚皮苷具有降血脂、镇静、抗氧化、抗肿瘤、抗真菌、抗动脉粥样硬化作用,推测这2 种物质可能是影响谷子养胃的重要代谢物。在栽培品种晋谷21 中,山奈酚3-O-芸香糖苷等黄酮类物质的含量显著低于农家种牛毛白。有研究表明将光敏核不育系 A2(PTGMS A2)分别与张杂谷 3、5、6、13 号、敦煌2 号杂交产生的后代叶片进行代谢组分析,发现叶片中含107 种黄酮类物质且过量积累,主要以糖基化形式存在。不同谷子品种杂交后对其后代进行综合代谢分析,将有助于开发杂交表现的预测模型,并实现谷子的遗传育种目标[21]。鉴于山奈酚3-O-芸香糖苷等黄酮类物质重要的药用功能,在未来分子品质育种中,可通过适当提高该物质在栽培品种中的含量,提高谷子中的功能性成分,为谷子功能农业与发展提供参考依据。

图7 山奈酚3-O-芸香糖苷的代谢图和热图Fig.7 Metabolism and heatmap of nicotiflorin
猜你喜欢
走向世界(2024年9期)2024-03-19 13:58:41
世界科学技术-中医药现代化(2021年10期)2021-03-02 05:52:00
核农学报(2020年4期)2020-03-13 14:19:42
山西农业科学(2019年10期)2019-10-25 02:28:38
阅读(低年级)(2019年5期)2019-07-18 19:13:50
中成药(2018年9期)2018-10-09 07:18:46
牡丹(2018年31期)2018-01-03 12:33:26
今日农业(2017年4期)2017-12-22 11:32:22
作文评点报·作文素材小学版(2015年35期)2015-09-29 01:09:08
现代农业科技(2015年12期)2015-08-17 16:23:12
