
高光效种植模式下玉米间作平菇对玉米穗位叶光合特性的影响
2020-07-01 14:32:46杨小琴齐晓宁孙露莹张梦杰宋凤斌刘胜群李向楠朱先灿
土壤与作物 2020年2期
杨小琴,王 洋,齐晓宁,孙露莹,张梦杰,宋凤斌,刘胜群,李向楠,朱先灿
(1.中国科学院 东北地理与农业生态研究所,吉林 长春 130102;2.中国科学院大学,北京 100049)
0 引 言
间作通常指在同一块土地上种植两种及两种以上作物的种植模式,当一种作物生长到一定阶段,在其行间种植其它作物(一种或多种)被称为套种[1]。间套作的合理布局有利于高效利用光照、热量、水分及养分等资源[2],增加作物产量[3],提高土地当量比[4]。中国耕地面积仅占世界7%,却养活了世界人口的22%[5],多熟制种植下的间套作模式起着重要作用[6]。
玉米,世界三大粮食作物之一,是优质的饲料和工业原料[7]。世界玉米总产量的21.9%由我国贡献[8],而吉林省玉米产量占全国总产量的1/8[9]。长期以来,吉林省玉米种植以单作为主,土地利用效率不高,且黑土肥力逐渐下降[10]。平菇(Pleurotusostreatus),隶属于真菌界担子菌门,因其味道鲜美、营养成分含量高,逐渐成为一种重要的食用菌[11]。在荫蔽湿润环境条件下,平菇生长旺盛,会产生大量菌丝体和非特异性的木质纤维素酶[12],因而常作为间作作物以促进养分等资源的有效利用[13]。蚕豆间作蘑菇研究显示,与单作相比,蚕豆间作牡蛎蘑菇增加了蚕豆干种子产量和牡蛎蘑菇担子菌产量[14]。已有研究表明,玉米间作大豆[15]、花生[16[、苜蓿[17]及小麦[18]等系统中,田间作物高矮相间,光照条件有效改善,玉米功能叶的光合速率提高,间作处理下玉米产量均明显高于单作。
从20世纪90年代起,玉米间作平菇的生产实践和试验研究顺利开展,间作条件下玉米和平菇亩产提高,经济效益良好[19]。但玉米平菇间作种植模式的研究大多偏重技术,基础研究不足,在玉米高光效种植模式下,对玉米平菇间作共生期内玉米穗位叶的光合作用及关键酶活性的研究较为缺乏。
玉米物质生产的基础很大程度来源于叶片的光合作用,在玉米生长中后期,穗位叶光合指标高低、干物质积累速率与分配情况对产量有重要影响[20]。另外,穗位叶光合作用强弱与其关键酶的活性密切相关[21]。磷酸烯醇式丙酮酸羧化酶(PEPC)是C4植物光合碳同化途径的第一个关键酶[22];苹果酸脱氢酶(MDH)可以催化苹果酸与草酰乙酸间的可逆转化,而苹果酸酶(NADP-ME)主要功能是催化苹果酸氧化脱羧生成CO2和丙酮酸,之后CO2进入卡尔文循环[23];核酮糖-1,5-二磷酸羧化酶(Rubisco)通过对碳固定以及光呼吸碳氧化的控制,对植物光合速率具有重要影响[24-25];同时,丙酮酸磷酸双激酶(PPDK)作为C4循环中CO2固定的主要限速酶,催ATP及丙酮酸生成磷酸烯醇式丙酮酸(PEP),为C4植物光合作用提供重要碳源[26]。
玉米高光效种植模式主要根据玉米株高、经纬度、北半球夏季太阳高度角及方位角,确定了吉林省中部地区玉米最佳种植垄距为160 cm+40 cm的组合垄(2行玉米播种在40 cm的窄行上,160 cm宽行为休闲行),垄向为磁南偏西20°,改善了光照在玉米群体内的分布,提高了垄间的通风透光能力[27],同时采用留高茬秸秆全株还田与休耕轮作,促进了有机质积累和耕地的可持续利用[28]。
本研究在玉米高光效种植模式的宽行(160 cm)间作平菇,以单作玉米为对照,主要探讨共生期内间作对群体微气象环境、玉米穗位叶光合作用及关键酶活性的影响,这有助于提高米菇间作体系下玉米叶片光合碳同化方面的认识,并为米菇间作生产实践提供一些理论支撑。
1 材料与方法
1.1 供试材料与试验设计
试验在长春综合农业试验站(125°24′E,43°54′N)连续开展2年(2017年、2018年),试验区土壤类型为典型薄层黑土,气候类型为中温带大陆性气候,年平均降雨量530.5 mm,年平均气温4.6 ℃,无霜期较短(138 d)。土壤的物理和化学性质如表1。

表1 试验地土壤理化性质
供试玉米和平菇分别为良玉99和紫孢侧耳。试验共设置3个处理(如图1),分别为:玉米平菇间作(CM)、单作玉米(C)和单作平菇(M)处理,重复3次,共9个小区(面积为20 m×8 m=160 m2),种植行向为磁南偏西20°。玉米平菇间作和单作玉米均采用高光效种植模式,即160 cm+40 cm组合垄,窄行(40 cm)上种植玉米,株距和种植密度分别为15 cm、6.5×104hm-2;玉米平菇间作处理布设方式如下:首先,在宽行(160 cm)内挖一条宽约40 cm、深约12 cm的畦沟(与玉米间距为27 cm);再者,将菌棒均匀地排布于畦沟内,并在其表面覆盖3 cm土;单作平菇处理的行距为160 cm,与间作相同,且平菇生长期内,采用间断性喷灌以保持土壤湿润。基施600 kg·hm-2的控释复合肥(总养分≥53%)。2017年,玉米播种和收获日期分别为4月26日、9月30日;在玉米抽穗期(7月12日)种植平菇,一茬菇、二茬菇收获日期分别为8月5日、9月3日。2018年,玉米播种和收获日期分别为4月27日、9月28日;7月11日种植平菇,一茬菇、二茬菇收获日期分别为7月21日、8月11日。
1.2 测定项目与方法
1.2.1 环境参数。在田间安置环境记录仪(邯郸市邯山瑞华电子有限公司)监测空气温湿度、CO2以及土壤温度。其中测定空气温湿度、CO2浓度探测器置于冠层内部(距离地面20 cm),土壤湿度探测器置于土壤内部(埋藏深度15 cm)。
1.2.2 光合荧光参数测定。2018年,在平菇种植10 d(一茬菇:7月21日)、20 d(无菇期:8月1日)和30 d(二茬菇:8月11日)后。用LI-6400型光合仪(美国LI-COR公司)测定玉米穗位叶的光合参数,光强设定为2 000 μmol·m-2·s-1的,重复4次。
平菇种植10 d(一茬菇)后,用的MultispeQ(美国PhotosynQ公司)测定玉米穗位叶的荧光参数。
1.2.3 样品采集及测定。2018年7月21日(一茬菇)、8月2日(无菇期)和8月11日(二茬菇)采集玉米穗位叶(中部)样品,并避开主叶脉,4次重复。将样品放入液氮罐冷冻,采样结束后,置于实验室-80 ℃冰箱保存。干样采集方法:用剪刀取玉米棒三叶(穗位叶及其上、下两叶),4次重复,放入干净的档案袋中保存,烘箱内105 ℃杀青30 min,75 ℃烘干,粉碎后测定可溶性糖和淀粉。
5种关键酶(PPDK、Rubisco、NADP-ME、MDH和PEPC)活性测定方法:取适量鲜样放入研钵,添加少许液氮后充分研磨,取0.1 g样品置于2 mL离心管内,添加0.9 mL磷酸缓冲盐溶液(0.01M pH 7.4),用混匀器将样品和溶液混合均匀,4 ℃下离心15 min,转速为4 000 rpm,取上清液待用。用酶联免疫法(ELISA)试剂盒(江苏酶标有限责任公司)测定5种关键酶活性。
考马斯亮蓝法测定可溶性蛋白质含量[29]:其原理为蛋白质与考马斯亮蓝G-250结合后颜色由红转蓝,会导致最大光吸收从460 nm变为595 nm,用分光光度计测定595 nm光吸收的增加量,最终可计算出可溶性蛋白质含量。
蒽酮法测定可溶性糖含量[29]:在浓硫酸作用下,经过脱水反应的糖类会产生蓝绿色的糠醛衍生物,其颜色深浅与糖含量成正比例关系。加入蒽酮试剂后,糖类与其反应生成的有色物质,用分光光度计可测定620 nm的吸收峰,计算后得到可溶性糖含量。
硫酸-苯酚法测定淀粉含量:在硫酸的作用下,多糖会水解成单糖后迅速脱水产生糖醛衍生物;加入苯酚试剂后,与其生成橙黄色化合物,用比色法测定后计算得出样品中的淀粉含量。具体操作:由于水解液(1 mL)样品浓度过高,加蒸馏水稀释100倍,取稀释后2 mL样液,加入1 mL苯酚(6%)溶液和5 mL浓硫酸,用混匀器将样品和溶液混合均匀后,放置20 min,测定490 nm的吸光度计算得到总还原糖量含量,该值乘以0.9即为淀粉含量。
1.2.4 收获测产。于2017-2018年玉米成熟期,取两处理具有代表性的20 m2果穗,测量其穗重,并取20个果穗标准样本,风干后测定出籽率和含水率,最终计算玉米产量。
1.2.5 土地当量比。应用土地当量比(LER)作为衡量间作产量优势的指标[30]。
LER=(Yip/Yp)+(Yim/Ym)
式中:Yip和Yim分别代表间作总面积上平菇和玉米的产量;Yp和Ym分别为单作平菇和单作玉米的产量。当LER>1为间作优势,当LER<1为间作劣势。
1.3 数据统计分析与作图
用Microsoft Excel 2007和SPSS 23进行数据的统计分析,用Origin 9.1作图。
2 结果与分析
2.1 玉米平菇间作对微环境参数的影响
温湿度和CO2浓度(外部环境条件)会影响玉米光合作用,其自身也受种植方式的影响。图2显示,玉米平菇间作共生期内(7月11日-8月23日),间作处理下空气湿度和CO2浓度为86.9%、504.8 ppm,显著高于单作玉米(26.8%、394.8 ppm)(P<0.01)。间作处理下空气温度和土壤温度为24.7 ℃、22.9 ℃,与单作玉米(24.0 ℃、23.2 ℃)相比无显著差异;无菇期(8月2日),间作的空气湿度和CO2浓度出现下降趋势,与单作玉米环境参数差异明显减小。
2.2 玉米平菇间作对玉米穗位叶的光合特性
单作与间作玉米穗位叶的净光合速率(PN)、胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr),随着时间有先上升后下降的趋势(图3)。间作玉米穗位叶PN在一茬菇(7月21日)和二茬菇(8月11日)分别为43.6 μmol CO2·m-2·s-1、38.8 μmol CO2·m-2·s-1,比单作高15.6%和25.7%,且在二茬菇差异达到显著(P<0.05),无菇期(8月1日)间作与单作玉米的PN无显著差异。一茬菇(7月21日)和二茬菇(8月11日)期间,间作玉米穗位叶的Gs分别为0.3 mol H2O·m-2·s-1、 0.2 mol H2O·m-2·s-1,均比单作高0.2 mol H2O·m-2·s-1;而在无菇期(8月1日)则显著低于单作玉米(P<0.05),Tr与Gs的变化趋势相同。
单作与间作玉米穗位叶的Ci无显著差异,无菇期(8月1日)间作的Ci为194 μmol CO2·m-2·s-1,低于单作玉米(232 μmol CO2·m-2·s-1),但一茬菇(7月21日)、二茬菇(8月11日)均高于单作玉米(P>0.05)。结果显示,间作处理下盛菇期提高了玉米穗位叶的PN,这将有助于玉米光合碳同化效率的提升。
2.3 间作玉米穗位叶的叶绿素荧光动力学参数
光合作用的机理可以通过叶绿素荧光参数来反映[31],叶绿素荧光技术是研究PSII功能和反应的有效手段[32-33]。最大光化学效率(Fv/Fm)和实际光化学效率(Y(Ⅱ)),分别指示植物的潜在最大光合能力(光合效率)和实际光合能力;PSII处调节性能量耗散的量子产量(Y(NPQ))和非调节性能量耗散的量子产量(Y(NQ))分别为光保护和光损伤的重要指标;如果 Y(NPQ)较高,则表示植物吸收了过剩的光能,但植物可以通过适当的调节(如热耗散)来保护自身;如果Y(NQ)较高,则表示保护性调节机制不能将植物接受的过剩光能完全消耗掉。
研究表明,一茬菇(7月21日),米菇间作模式下的玉米穗位叶Fv/Fm显著高于单作玉米(表2,P<0.05);间作玉米的Y(II)和Y(NQ)高于单作玉米,均未达到显著水平;而间作处理的Y(NPQ)显著低于单作玉米(P<0.05)。由此推测,米菇间作期(一茬菇)玉米穗位叶最大光合速率和实际光合速率的提高与光能传递、转化密切相关。

表2 间作对玉米荧光参数的影响
注:不同小写字母表示处理间在P<0.05水平上差异显著。
Note:Different samll letters indicate significant differences between treatments at 0.05 level.
2.4 间作玉米穗位叶的光合作用5种关键酶活性
间作对玉米光合作用5种关键酶的影响见图4,与单作玉米相比,玉米平菇间作一茬菇(7/21)时期,5种关键酶的活性差异均不显著;无菇期(8/2),除Rubisco酶活性稍高外,MDH、NADP-ME、PEPC、PPDK酶活性均低于单作玉米,但都未达到显著水平;而二茬菇(8月11日)间作提高了玉米穗位叶的MDH、NADP-ME、PEPC、PPDK、Rubisco酶活性,其中MDH酶活性显著高于单作玉米(P<0.01),是单作玉米的2.4倍,PEPC酶活性是单作玉米的1.7倍。
检验结果表明,生育时期对玉米的MDH(P<0.01)、NADP-ME(P<0.01)、Rubisco酶活性(P<0.01)有极显著影响(表3),种植模式对玉米5种光合作用关键酶的影响均不显著;但是,时期与种植方式的交互作用对MDH酶活性有极显著影响(P<0.01)。这表明,玉米间作平菇二茬菇时期提高了碳素同化的关键酶活性,为玉米生长中后期(灌浆成熟期)光合产物的积累奠定了基础。
2.5 间作玉米穗位叶的可溶性蛋白质、可溶性糖和淀粉
可溶性蛋白质(包含多种酶源、酶分子以及代谢调节物),在植物的蛋白质组分中扮演着最活跃的角色。间作处理下玉米棒三叶的可溶性蛋白质含量高于单作,在二茬菇(8月11日)期较为明显(图5,P>0.05);另外,生育时期对可溶性蛋白含量的影响显著(P<0.01)。与单作玉米相比,间作处理玉米棒三叶的可溶性糖及淀粉含量稍低(一茬菇和无菇期),但二茬菇稍高(P>0.05)。由此推测,玉米生育种中后期(二茬菇),间作处理下玉米有较充足的物质源(主要包括可溶性蛋白质、可溶性糖和淀粉等)为籽粒灌浆奠定基础。
2.6 玉米平菇间作对玉米产量和土地当量的影响
与单作玉米相比,2017年、2018年间作玉米百粒重分别增加了20.8%、3%,穗粒重增加了3.7%、2.3%(表4)。两年间,玉米净增产量为376 kg·hm-2、325 kg·hm-2,同时,平菇净增产量为16 560 kg·hm-2、14 064 kg·hm-2,LER均大于1,分别为2.5和2.4,间作产量优势明显。

表3 不同种植模式、时期及其相互作用对玉米光合作用5种关键酶的影响
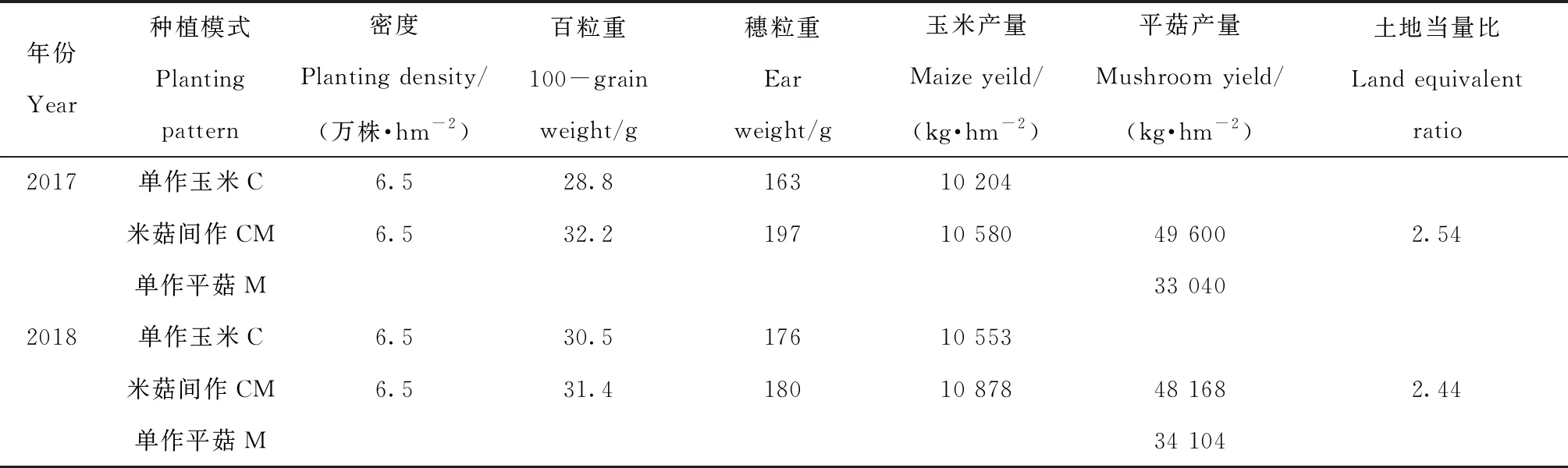
表4 玉米产量构成因素及土地当量比
3 讨论与结论
光合作用合成有机物,是作物产量形成的基础[34-36],作物光合作用能力强弱通常可以用净光合速率(PN)来衡量。CO2是作物进行光合作用的重要原料,有研究显示作物光合作用会随CO2浓度(不超过500 μmol·mol-1)的升高而增强[37]。研究棉田套种平菇、玉米间作平菇试验表明,间作系统有助于调节光照、温度及水分等资源[38],平菇出菇良好,并释放出大量的CO2,为棉株和玉米光合作用提供较为充足的原料,并提高了光合效率,促进棉花和玉米生育后期生物量的累积[39-40]。本研究表明,一茬菇期,间作体系下玉米穗位叶最大光化学效率(Fv/Fm)和PN较单作分别提高了4%和15.6%,二茬菇期,间作玉米PN比单作提高了25.7%,这可能是因为间作期内的田间温湿度适宜,其中空气温度、空气湿度分别比单作玉米提高了3.6%和9.1%,同时CO2浓度增加了22.5%,玉米光合作用可利用碳源增加,促使玉米穗位叶的光合速率提高,籽粒灌浆期间将有更多的光合物质向籽粒运输和分配。无菇期,间作体系下的空气温湿度和CO2浓度逐渐下降,与单作玉米环境条件趋于一致,且间作玉米穗位叶的Fv/Fm、PN较单作无明显差异。另外,与单作玉米相比,间作处理增加了玉米和平菇产量,土地当量比提高,这与很多间作系统研究[15,17,41]的结论相符。
玉米穗位叶的Rubisco、PEPC、MDH、NADP-ME和PPDK等酶活性,在一定程度上反映了碳素同化的效率高低[42]。研究表明,玉米抽丝期(无菇期)-灌浆期(二茬菇)穗位叶的Rubisco、NADP-ME和PPDK酶活性显著降低,这与春玉米吐丝期-灌浆期-成熟期光合作用Rubisco等酶活性呈现逐渐降低变化趋势的研究一致[43]。
植物生长季,Rubisco的羧化效率提高(5%~25%),将促进叶片的光合速率,并可能使干物质产量增加几倍[44]。研究表明,二茬菇期,间作的Rubisco酶活性较单作提高13%,可能促进玉米叶片光合作用并有利于干物质积累。与单作玉米相比,二茬菇间作处理的穗位叶MDH酶活性显著提高(P<0.05),且NADP-ME、PEPC和PPDK酶活性分别提高了12%、74%和14%。同时,间作处理平菇生长释放大量的CO2,若玉米叶片有效吸收,PEPC酶同化CO2生成更多的草酰乙酸,原料的增加和MDH酶活性的显著提高,将催化较多的草酰乙酸转化为苹果酸,而苹果酸经NADP-ME酶氧化脱羧生成较为充足的丙酮酸和CO2,而Rubisco酶活性提高对CO2同化过程中起关键作用,这一过程可能会有效调节玉米穗位叶的光合速率,积累较多的光合产物,为生育中后期玉米籽粒灌浆奠定基础[45]。但也有研究发现,活化的Rubisco量(而不是Rubisco的总量)会影响植物光合速率[46],主要原因是钝化态Rubisco的羧化/加氧活性需要通过Rubisco活化酶调节[47]。无菇期,间作处理光合速率比单作低,可能是由于间作玉米活化的Rubisco量较少,以及MDH、NADP-ME、PEPC、PPDK酶活性较低导致的。
综上,在高光效种植模式下玉米间作平菇共生期内,田间作物群体内空气湿度和CO2浓度显著提高,在平菇生长旺盛期(8月11日),间作玉米的净光合速率、5种关键酶活性、3种光合产物都高于单作玉米,这为玉米生长提供了更为适宜的生长环境,并为其光合作用提供较为充足的碳源,促进了玉米生长中后期(灌浆期-成熟期)光合效率的提高和产物的累积,玉米产量增加,土地当量比提高。这将对吉林省中部地区玉米间作平菇的高产栽培提供一些理论支持。玉米间作平菇系统显著改善了作物群体微环境条件,可能会对不同层位玉米叶片产生影响,另外,平菇(真菌界担子菌门)与根际土壤微生物相互作用和物质能量交换,这都会影响玉米光合产物积累与分配,因而都值得进一步深入研究。
猜你喜欢
故事作文·低年级(2023年1期)2023-02-23 07:19:22
作物学报(2022年8期)2022-05-29 06:22:44
广西林业科学(2022年2期)2022-05-09 07:49:54
作物杂志(2022年6期)2022-02-03 04:56:06
小学生(看图说画)(2020年10期)2020-10-29 01:44:32
湖南农业大学学报(自然科学版)(2020年4期)2020-08-28 01:43:28
山东农业科学(2019年11期)2019-12-24 01:11:27
江苏农业科学(2019年11期)2019-07-22 01:32:45
山西农业大学学报(自然科学版)(2018年12期)2018-12-04 08:18:34
干旱地区农业研究(2017年5期)2017-12-18 06:18:18
