
环境因子对大兴安岭天然落叶松次生林主要树种更新的影响1)
2020-06-27 16:08:54祝子枭刘兆刚董灵波陈莹张凌宇由珈齐
东北林业大学学报 2020年6期
祝子枭 刘兆刚 董灵波 陈莹 张凌宇 由珈齐
(东北林业大学,哈尔滨,150040)
森林天然更新,是利用林木自身繁殖和恢复能力,在林地或林迹地上形成新一代幼林的过程,是森林生态系统中资源的再生产[1]。天然更新,从种子产生、扩散、萌发、幼苗定居和建成到衰老枯倒,每一个阶段都受到各种环境因子的影响[2]。影响更新因素的研究,主要集中在母树与种源[3-5]、林分因子[6-9]、土壤因子[10-11]、气候因子[12-13]、干扰因子[14-17]等几方面;对土壤因子方面,并未考虑不同土壤厚度、理化性质对更新的影响。更新苗在各个生长阶段,根系扎根深度不同,直接影响林分的更新。按高度级对更新苗进行等级划分,但是高度划分并未考虑不同树种之间的更新差异。国内对更新的影响因子分析方法,主要有相关性分析、主成分分析、多元回归统计分析、方差分析、灰色关联分析等[18-21]。冗余分析是生态学领域一种常见的分析方法,在林分更新方面应用的较少,本文引入冗余分析法探讨林分更新的影响因子。
落叶松(Larixgmelinii)林是大兴安岭地区的顶级群落之一,是大兴安岭地区分布面积最大、分布最广的基本森林类型[22-23],在海拔300~1 200 m均有分布。天然林主要分布在海拔500 m以上的山区,由于人为破坏严重,现有的落叶松林多属于次生林。本文研究多种因素对大兴安岭天然落叶松林更新的影响,了解森林更新与环境因素之间的关系,旨在为科学实施森林可持续经营管理、实现人工林经营过程的近自然化提高参考。
1 研究区概况
研究区位于大兴安岭地区翠岗林场(123°41′~125°25′E,51°21′~52°10′N)、新林林场(123°41′~125°25′E,51°20′~52°10′N)、壮志林场(123°29′~124°18′E,50°59′~51°24′N)。大兴安岭地区温度变化较大,冬季寒冷漫长,达7个月之久,昼短夜长,日照少,平均气温-28 ℃;冬季降水量较少,仅占年降水量的10%。夏季时间6—8月份,日照时间较长,可达17 h,昼夜温差大;夏季由于东南季风气候的影响,降水多,主要集中在7—9月份。由于降水量的差异,大兴安岭冬季干燥,而冬季气候湿润温和,初霜、终霜时间都较早。主要林分类型为白桦林、落叶松林、落叶松与白桦为主的针阔混交林。主要树种有落叶松(Larixgmelinii)、白桦(Betulaplatyphylla)、云杉(Piceaasperata)、樟子松(Pinussylvestrisvar.mongolicaLitv)、山杨(Populusdavidiana)等;灌木层树种有笃斯越桔(Vacciniumuliginosum)、杜香(Ledumpalustre)、兴安杜鹃(Rhododendrondauricum);地表草本有蕨类(Adiantumspp.)、莎草(Cyperusspp.)、舞鹤草(Maianthemumdilatatum)、地榆(Sanguisorbaofficinalis)等。
2 研究方法
2.1 样地设置
本文以2018、2019年,在大兴安岭地区翠岗林场、新林林场、壮志林场设定的57块固定样地作为原始数据(见表1),其中:翠岗林场10块0.06 hm2、10块0.1 hm2固定样地,新林林场18块0.1 hm2固定样地,壮志林场10块0.1 hm2、9块0.08 hm2固定样地。调查并记录乔木层(胸径≥5 cm)每木树种、状态、胸径、树高、第一活枝高、第一死枝高、位置坐标(x、y)、东南西北4个方向的冠幅等信息;调查并记录更新层(胸径<5.0 cm)每木树种、状态、地径、胸径、树高、位置坐标(x、y)、更新方式(实生和萌生)等。每个样地的四角点,分别设置1个2 m×2 m的灌木调查样方、1 m×1 m的草本调查样方,调查每个小样方中灌木和草本的种类、平均高、盖度等信息,并记录样地的海拔、坡向、坡位、坡度、土壤类型等信息。
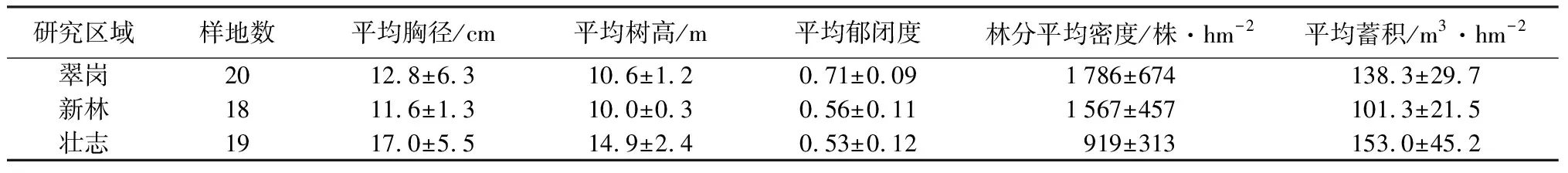
表1 样地概况
土壤样品采集:在每个样地沿样地对角线取中心及两样方角的土壤样品,分别取A0层(枯枝落叶层,分解、半分解的有机物质积累的层次,木本植被下的森林土壤最为明显)、A1层(腐殖质层,由于腐殖质的累积,腐殖质和矿质养料含量丰富,且结合紧密,多呈良好的团粒结构,土色较深)、A2层(淋溶层,由于雨水的淋洗作用,土体中易溶性盐类及铁、铝水化物、腐殖质胶体受到淋失,向下移动,使该层腐殖质及养分含量减少,土色较浅)土壤,荫干后置于密封袋中,带回实验室荫干后参照《土壤农业化学分析方法》测定其化学性质,测量并记录A0、A1、A2层的厚度[24-25]。
2.2 更新幼苗等级划分
结合更新苗等级划分标准(幼苗,株高(h)<30 cm;小苗,30 cm≤h<130 cm;幼树,h≥130 cm或胸径≤5 cm),将更新苗按照株高划分为3个等级,即幼苗、幼树、小树。按照落叶松、白桦、不分树种3种情况,对每个样地更新幼苗分等级计数,分别代表各个样地主要树种不同生长阶段的更新情况,从而对天然落叶松次生林更新情况进行分析。其中,更新密度划分为6个类别:落叶松幼苗、白桦幼苗、落叶松小苗、白桦小苗、落叶松幼树、白桦幼树。
2.3 环境因子的选取
本文选取25个环境因子(见表2),以诠释地形、林分条件、空间结构、树种组成、土壤环境等方面的环境变化,通过尽可能多的环境因子探讨其对天然落叶松次生林更新的影响。将25个环境因子划分为2类,即林分因子12个指标、土壤因子13个指标。土壤样品分为枯枝落叶层、腐殖质层、淋溶层3层,对腐殖质层、淋溶层分别测量其化学性质,包括pH值、有机质、全氮、全磷、全钾指标。
2.4 非对称约束排序
冗余分析(RDA),是一种将回归分析法与主成分分析法融合进行排序的分析方法,也是对多响应变量回归分析的延伸拓展。冗余分析是响应变量矩阵与解释变量矩阵之间,多元多重线性回归拟合矩阵的主成分分析。冗余分析是生态学领域常用分析方法,Legender和Gallagher开发了一系列物种转化数据,使物种数据适用于冗余分析,此后,冗余分析得到了更加广泛的应用[26]。
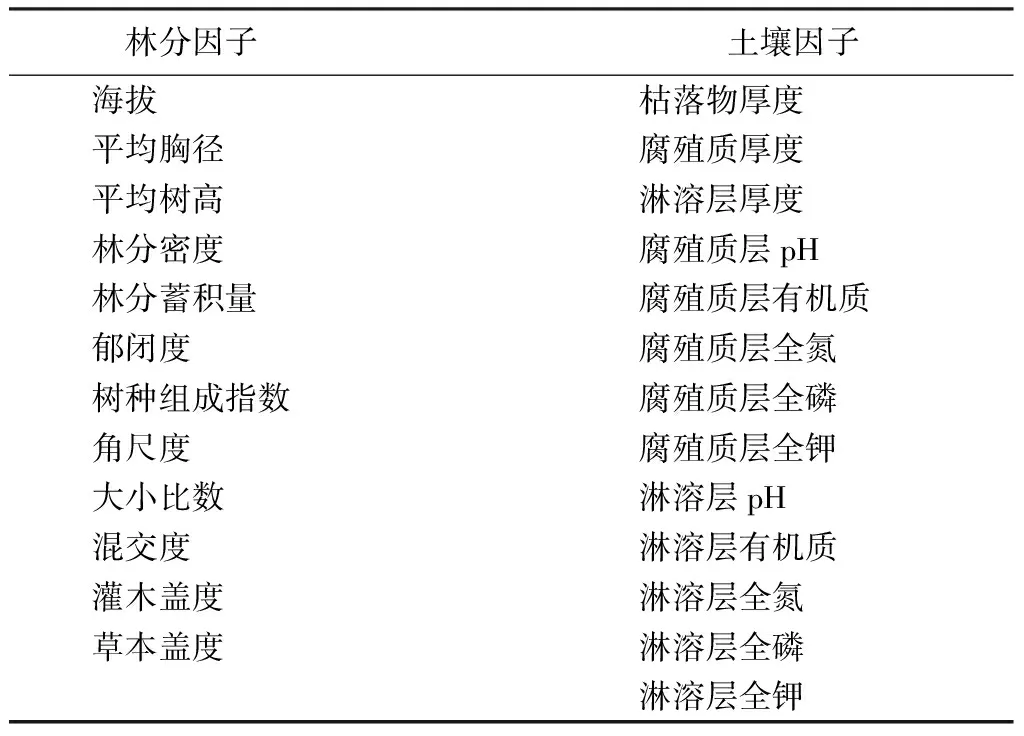
表2 冗余分析使用的生境因子
为了寻找简约模型,解决解释变量之间的共线性问题,本文采用解释变量的前向选择。变量的共线性程度可以用方差膨胀因子(FVI)度量,当FVI大于20时,说明与其他解释变量之间存在严重的共线性问题。
变差分解,即量化两组或多组变量单独及共同解释的变差。1992年,Borcard et al.[27]首先提出变差分解的概念,并且发现其分解过程,这对于多元生态数据分析有重要的意义。Peres-Neto et al.[28]提出使用校正决定系数(R2)促进变差分解的使用。通过变差分解,将解释变量分为3部分,即林分变量解释部分、土壤变量解释部分、共同解释部分。
3 结果与分析
3.1 冗余分析
对物种数据进行趋势对应分析(DCA),获得物种数据4个排序轴,其中最长轴梯度为2.1109。如果最长排序轴小于3.0,排序分析方法选择应用线性模型的冗余分析更合理[29]。
为了确认不同区域之间的是否存在显著差异,运用组合聚类分析与排序分析的方法,在冗余分析排序图基础上添加聚类树(见图1)。3组聚类簇均包含3个区域的样地,并且3组聚类簇的样方点在排序坐标上分布比较均匀,说明大兴安岭这3个区域无显著性差异。
冗余分析结果显示,25个环境因子对林分更新密度的贡献率为77.16%(见表3),利用蒙特卡罗(Monte Carlo)检验(置换次数为999),全模型前两个排序轴显著(p<0.05)。前4轴累计贡献率为97.33%,前2轴累计贡献率为85.63%,因此,可以用冗余分析前2个排序轴反映大兴安岭天然落叶松林的更新状况(见表4)。第1排序轴主要反映了林分平均胸径、平均树高、林分蓄积、草本盖度、林分密度、枯落物厚度、腐殖质层有机质的影响;第2排序轴主要反映了海拔、林分蓄积、林分郁闭度、树种组成、角尺度、腐殖质层全磷质量分数的影响。
表3冗余分析前4个排序轴的累计贡献率
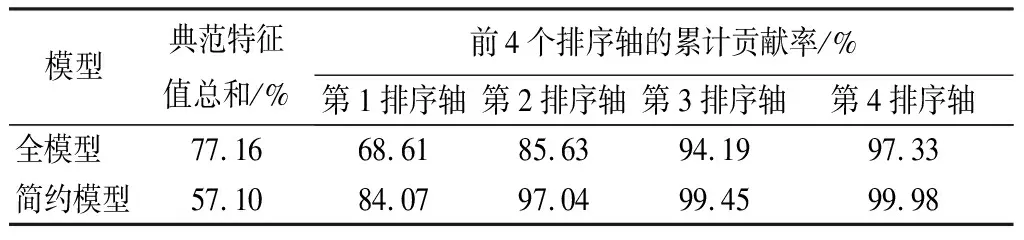
模型典范特征值总和/%前4个排序轴的累计贡献率/%第1排序轴第2排序轴第3排序轴第4排序轴全模型77.1668.6185.6394.1997.33简约模型57.1084.0797.0499.4599.98
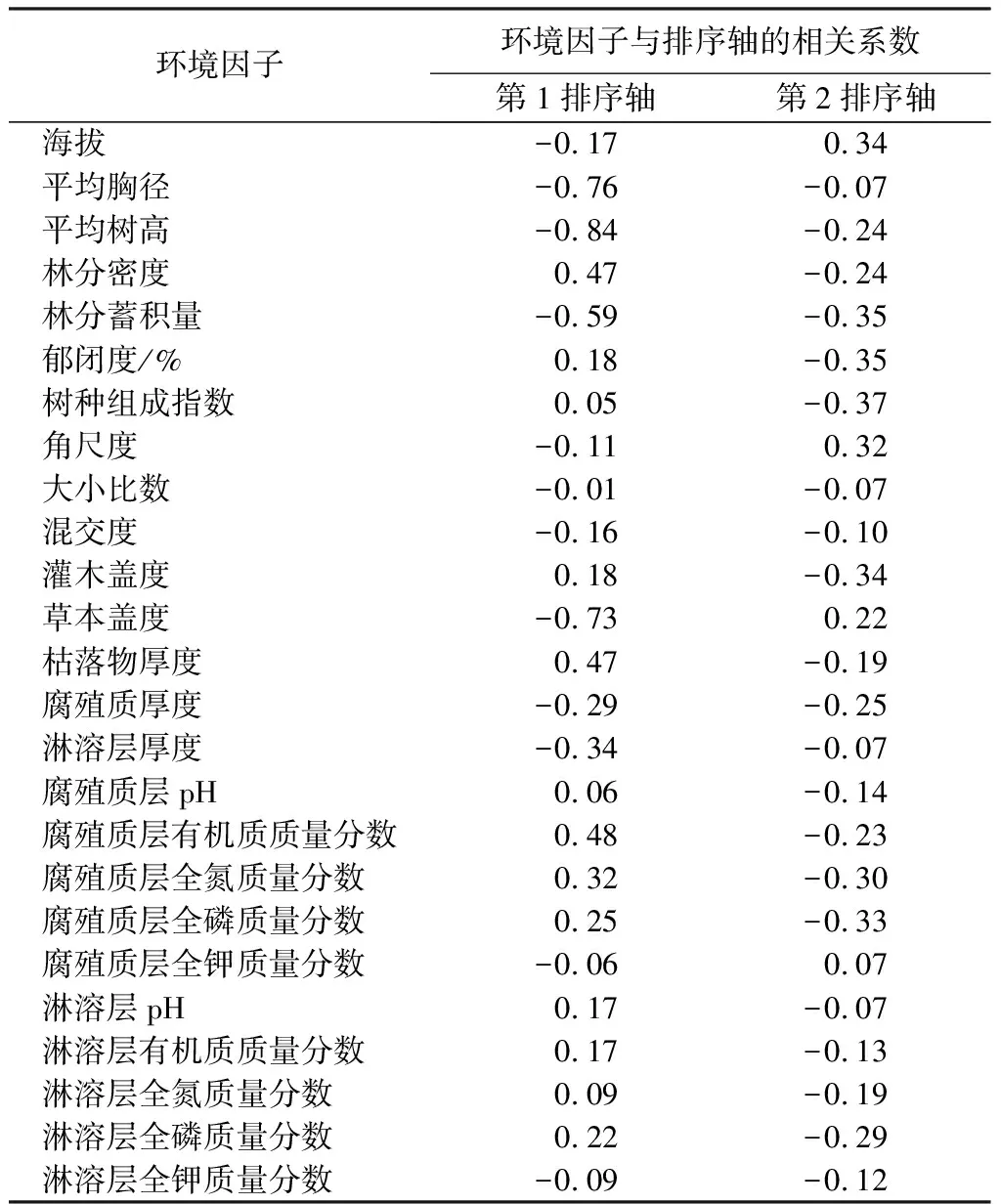
表4 环境因子与排序轴的相关关系
更新林分可以分为两组:第一组为落叶松幼苗、白桦幼苗、落叶松小苗,3者聚集于坐标原点附近,表明这些林分在整个区域分布均匀或者分布于环境梯度的中值区域;第二组为白桦小苗、落叶松幼树、白桦幼树,3者箭头比较长,比第一组分布较散,表明这些林分受个别环境因子影响较大(见图2)。
图面中的数字1~57为样地编号。ELE为海拔;DBH为平均胸径;H为平均树高;N为林分密度;M为林分蓄积量;CW为郁闭度;SCI为树种组成指数;AS为角尺度;SR为大小比数:MA为混角度;SC为灌木盖度;HC为草本盖度;LTH为枯落物厚度;HTH为腐殖质厚度;STH为淋溶层厚度;PH1为腐殖质层pH;Oma1为腐殖质层有机质质量分数;TN1为腐殖质层全氮质量分数;TP1为腐殖质层全磷质量分数;TK1为腐殖质层全钾质量分数;PH2为淋溶层pH;Oma2为淋溶层有机质质量分数;TN2为淋溶层全氮质量分数;TP2为腐殖质层全磷质量分数;TK2为淋溶层全钾质量分数;ND1为落叶松幼苗;ND2为白桦幼苗;ND3为落叶松小苗;ND4为白桦小苗;ND5为落叶松幼树;ND6为白桦幼树。
图2环境因子对林分更新密度影响在冗余分析中的排序
全模型虽然能较大程度反映落叶松林更新苗木间的显著差异,但是环境因子较多,大部分环境因子对更新状态的影响很小,而且不利于后续分析。因此,采用解释变量的前向选择法进行变量筛选,寻找冗余分析简约模型。简约冗余分析结果显示,枯落物厚度、腐殖质层全氮质量分数、郁闭度、草本层盖度、林分平均5个因子对林分更新密度影响最大,因此这5个因子进入简约模型(见图3)。这5个环境因子(方差膨胀因子均小于2.5)对林分更新密度影响的累计贡献率为57.1%,占25个环境因子对林分更新密度贡献率的74%,前2个排序轴的累计贡献率为97.04%(见表3)。
3.2 简约模型的林分更新密度等值线
为了更加直观的分析环境因子对林分更新密度的影响,提取冗余分析简约模型中的变量,即林分平均高、林分郁闭度、枯落物厚度、草本盖度、腐殖质层全氮质量分数5个环境因子。作出不同树种、不同等级更新苗密度的冗余分析等值线图(见图4~图9),这5个环境因子对林分更新密度影响的贡献率为57.1%,占全部环境因子对林分更新密度影响贡献率的74%。
由图4可见:落叶松幼苗更新密度等值线形成1个半封闭圆弧,涵盖了枯落物厚度、腐殖质层全氮质量分数、郁闭度3个因子箭头经过区域,因而落叶松幼苗更新密度与这3个因子相关性不明显。在垂直于落叶松幼苗更新密度等值线上,随着平均高、草本盖度增大,更新层密度逐渐减小。等值线密度整体变化很小,更新密度多为10株/hm2,说明落叶松更新幼苗稀少。
图面中的数字1~57为样地编号。LTH为枯落物厚度;TN1为腐殖质层全氮质量分数;CW为郁闭度;HC为草本盖度;H为平均树高;ND1为落叶松幼苗;ND2为白桦幼苗;ND3为落叶松小苗;ND4为白桦小苗;ND5为落叶松幼树;ND6为白桦幼树。
图35个环境因子对林分更新密度影响在冗余分析中的排序
图面中的数字1~57为样地编号。LTH为枯落物厚度;TN1为腐殖质层全氮质量分数;CW为郁闭度;HC为草本盖度;H为平均树高。
图4落叶松幼苗更新密度等值线
由图5可见:白桦幼苗更新密度等值线,在坐标原点呈椭圆,外围呈半封闭圆弧,而椭圆等值线为零值,说明草本盖度、林分平均高变化对白桦幼苗更新密度影响不显著。在垂直于白桦幼苗更新密度等值线上,随着枯落物厚度、腐殖质层全氮质量分数、郁闭度的增大,白桦更新幼苗密度增大。除了44、45、55、56、57号样地外,其余样地基本无白桦幼苗更新,说明翠岗、新林地区样地白桦幼苗更新不良;壮志地区部分样地虽然有白桦更新幼苗,但其更新密度也较低。
图面中的数字1~57为样地编号。LTH为枯落物厚度;TN1为腐殖质层全氮质量分数;CW为郁闭度;HC为草本盖度;H为平均树高。
图5白桦幼苗更新密度等值线
由图6可见:在垂直于落叶松小苗更新密度等值线上,更新密度从左至右逐渐增大;即更新密度,与枯落物厚度、腐殖质层全氮质量分数、郁闭度3个因子呈正相关,与草本盖度、林分平均高呈负相关。
图面中的数字1~57为样地编号。LTH为枯落物厚度;TN1为腐殖质层全氮质量分数;CW为郁闭度;HC为草本盖度;H为平均树高。
图6落叶松小苗更新密度等值线
由图7可见:在垂直于白桦小苗更新密度等值线上,随着草本盖度、林分平均树高的增大,白桦小苗更新密度减小。随着枯落物厚度、郁闭度、腐殖质层全氮质量分数的增大,更新密度增大。从左至右,密度等值线逐渐变密,白桦小苗更新密度差异较大,大部分样地白桦小苗更新不良,缺乏更新。
由图8可见:沿着草本盖度、林分平均高箭头方向形成一层层半闭合圆弧;沿着水平轴方向等值线密度逐渐变小;在垂直于落叶松幼树更新密度等值线上,随着草本盖度、林分平均高的增加,落叶松幼树更新密度先增加后减小。更新密度与枯落物厚度、腐殖质层全氮质量分数、郁闭度3个变量呈负相关。
由图9可见:在垂直于白桦幼树更新密度等值线上,随着草本盖度、林分平均高的增大,白桦幼树更新密度减小。随着枯落物厚度的增大,白桦幼树更新密度增大。腐殖质层全氮质量分数、郁闭度箭头与密度等值线基本保持平行,故腐殖质层全氮质量分数、郁闭度变化对白桦幼树更新密度影响不显著。
图面中的数字1~57为样地编号。LTH为枯落物厚度;TN1为腐殖质层全氮质量分数;CW为郁闭度;HC为草本盖度;H为平均树高。
图7白桦小苗更新密度等值线
图面中的数字1~57为样地编号。LTH为枯落物厚度;TN1为腐殖质层全氮质量分数;CW为郁闭度;HC为草本盖度;H为平均树高。
图8落叶松幼树更新密度等值线
图面中的数字1~57为样地编号。LTH为枯落物厚度;TN1为腐殖质层全氮质量分数;CW为郁闭度;HC为草本盖度;H为平均树高。
图9白桦幼树更新密度等值线
结合图4、图5分析,落叶松和白桦幼苗均存在更新不良的问题,绝大部分样地出现无更新幼苗的情况。结合图6、图7发现,白桦小苗更新密度整体好于落叶松小苗的更新密度;落叶松小苗更新密度各样地间差异较小,而白桦小苗更新密度等值线疏密变化较大,即各样地间更新差异较大。结合图8、图9分析,落叶松和白桦幼树更新密度各样地间相差不大,二者均存在更新不良的问题。
整体看,林分更新密度与林分平均高、草本盖度呈负相关,与枯落物厚度、腐殖质层全氮质量分数、郁闭度3个因子呈正相关。其中:落叶松幼树与其他苗木更新密度稍有差异,落叶松幼树更新密度与林分平均高、草本盖度相关性不呈线性增长趋势,呈先增大后减小的趋势;与枯落物厚度、腐殖质层全氮质量分数、郁闭度呈负相关。
3.3 环境因子对林分更新密度的影响
为了量化林分因子、土壤因子两组因子单独及共同影响的差异,本文采用变差分解的方法,分别对林分因子、土壤因子进行解释变量前向选择。其中,林分因子简约组合为林分平均高、草本盖度、郁闭度,土壤因子简约组合为枯落物厚度、淋溶层厚度、淋溶层全磷质量分数、腐殖质层厚度。林分因子单独对林分更新密度影响的贡献,占环境因子对林分更新密度影响总贡献率的72.28%;而土壤因子对林分更新密度影响贡献率较低,只占环境因子对林分更新密度影响总贡献率的13.14%;两组因子共同对林分更新密度影响的贡献率,占环境因子对林分更新密度影响总贡献率的14.58%(见图10)。总体看,林分因子对林分更新密度的影响,大于土壤因子对林分更新密度的影响。
4 讨论
张树梓等[30]研究表明,Ⅲ龄级林分天然更新苗在早期生长中,受土壤枯落物因子限制比较大,随着生长继续,林分结构因子逐渐代替土壤枯落物因子成为主要的更新限制因子,而这种现象在Ⅳ龄级林分中体现不明显。王笑梅等[31]研究发现,当冬青重要值较高时,林分结构因子是主要影响因子,土壤因子影响较小;当冬青重要值较低时,土壤因子的影响大于林分结构因子。而本文的变差分解结果表明,林分因子对林分更新密度的影响大于土壤因子。根据林分更新密度等值线图(见图4、图5)发现,大兴安岭地区更新幼苗低萌发量,导致在排序时,无法体现土壤因子对更新苗在前期生长的限制影响;而更新小苗、幼树的密度与更新幼苗密度相比,明显增大;因此结果显示,林分因子对天然更新的影响,大于土壤因子。此外,本文只选取了土壤因子6个指标,即全氮、全钾、全磷、pH值、有机质、土壤厚度,还不能全面反映林分的养分状况和物理性质。实际林分中,对更新产生影响的土壤因子还有很多,在以后的研究中,应对土壤的速效钾、有效磷、孔隙度、凋落物持水量等方面进一步研究,从而全面揭示土壤因子对更新的影响。
林分更新与环境因子的关系比较复杂,不同的研究结果不尽相同。灌草植被的地上、地下竞争,改变了林下光照、土壤湿度、温度以及凋落物的分布[9]。林下层对幼苗出现和定居的干预,不仅直接与其进行资源竞争,也间接影响种子捕食者行为,从而对幼苗定居与生长产生影响[32-33]。不同灌草的地上、地下竞争,会影响更新苗的光合作用与生物量的积累[34]。杂草的地上竞争加剧更新苗的空间竞争压力,并限制更新苗的光合作用;地下竞争又争夺土壤养分与水分,导致更新苗的光补偿点提高,净光合速率下降限制幼苗的生长发育。即随着林下草本盖度增加,更新密度呈下降趋势。
林地凋落物对种子萌发、幼苗建立和生长有双重作用[35]。有研究表明,枯落物层对种子存在自毒作用[36]。即使有充足的种源也很难形成更新,种子散落在枯落物层,林内湿度造成种子发霉,失去活力而无法萌发。枯落物覆盖的种子存活率明显高于裸露微生境,且与枯落物厚度呈正相关[37]。枯落物对种子有保护作用,防止动物觅食。此外,枯落物层导热不良,防止土壤升温过快而造成水分蒸腾,枯落物分解后还能为土壤提供营养元素。白桦种子和落叶松通常停留在枯落物层[38-39],这也验证了本文更新密度与枯落物呈正相关的结论。
土壤氮素质量分数是幼苗生长的限制因子之一[40]。宋新章[41]研究发现,全氮质量分数对幼苗的密度有显著的决定性作用,全磷质量分数对幼苗的径生长有较大的促进作用,全钾质量分数对幼苗的高生长有很强的促进作用。而本文也证明了更新密度与腐殖质层全氮质量分数呈正相关。
在郁闭度小于0.7时,白皮松林下天然更新密度较大;当郁闭度大于0.8时,天然更新密度小,而且成活率降低[42]。兴安落叶松与白桦皆为阳性树种,当林分郁闭度过高时,林下光照不足,将限制白桦与落叶松更新,大部分更新未进入主林层之前便逐渐枯死。但当郁闭度过小时,林下光照过强,土壤温度过高,水分流失较大,也不利于林分更新。而大兴安岭大部分林分郁闭度偏低,因而结果显示,林分更新密度与郁闭度呈正相关。
叶面积指数与林分平均高呈显著负相关[43]。叶面积指数越大,林分郁闭程度越差,即林分平均高与郁闭度呈负相关,更新密度与林分平均高呈负相关,本研究结果与其他学者研究结果一致[44]。
5 结论
大兴安林地区,主要影响林分更新的环境因子是:林分平均高、林分郁闭度、枯落物厚度、草本盖度、腐殖质层全氮质量分数,这5个环境因子对林分更新密度影响的贡献率为57.1%。从变量分解结果看,林分因子对林分更新密度的影响大于土壤因子。
大兴安岭地区主要树种更新不良,缺乏幼苗更新。3个区域的更新无明显差异,落叶松、白桦树种之间更新密度差异明显,幼苗、小苗、幼树3个等级更新密度差异明显。整体看,大兴安岭地区主要树种更新密度,与林分平均高、草本盖度呈负相关,与枯落物厚度、腐殖质层全氮质量分数、郁闭度呈正相关。
猜你喜欢
《学习方法报》小学语文四年级(2023年26期)2023-04-29 00:44:03
安徽文学(2023年4期)2023-04-11 22:49:27
福建林业(2020年5期)2020-03-18 08:23:02
文苑(2019年20期)2019-11-20 02:12:33
文苑·经典美文(2019年10期)2019-10-16 02:47:16
热带林业(2019年4期)2019-03-05 09:53:58
中国环境科学(2018年10期)2018-10-29 04:22:40
森林工程(2018年3期)2018-06-26 03:40:46
农民致富之友(2014年7期)2014-10-21 20:06:25
水土保持通报(2014年5期)2014-06-09 08:26:46
