
芥子碱和硫酸铜对芥子碱降解菌酶活力的影响
2020-06-22 08:36黄小丽邹成义
西南农业学报 2020年5期
余 丹,黄小丽,邓 卉,冯 杨,李 斌,邹成义 ,屈 东
(1.四川省畜牧科学研究院饲料研究所,四川 成都 610066;2.动物遗传育种四川省重点实验室,四川 成都 610066;3.四川农业大学动物科技学院,四川 成都 611130)
【研究意义】芥子碱是以菜籽粕为代表的十字花科类饲料资源的重要次生代谢产物[1],易被动物肠道微生物降解为腥味物质,诱发畜禽的鱼腥味综合征,严重影响畜产品品质和人类健康[2];它味道苦涩,是脱除了硫甙的菜籽产品适口性较差的主要原因,严重影响动物采食量[3];它可与蛋白质结合降低蛋白质的消化吸收率[4];芥子碱的抗雄激素活性能抑制精子成熟,引起动物生殖功能障碍[5]。随着双低菜籽粕的推广和应用,以芥子碱为代表的多酚类物质已成为菜籽粕最主要的抗营养因子[6]。因此,探索和研究芥子碱的降解和消减技术,不仅有利于十字花科类地方特色饲料资源的开发和利用,更有利于减少我国饲料工业对大豆粕等大宗饲料原料的依存度,为我国饲料工业稳定发展做出贡献。【前人研究进展】利用微生物发酵产生的酶蛋白降解芥子碱,是目前解决芥子碱抗营养问题最有效最安全的方法[7]。降解芥子碱的微生物包括真菌和细菌[8-9]。相对于真菌而言,细菌基因简单,新陈代谢旺盛,培养条件要求不高,能够在较短时间内进行大量繁殖,更有利于商业化生产。【本研究切入点】本项目组近年来专注于芥子碱及其降解菌的研究,通过以芥子碱为唯一碳源的培养基,已成功筛选出了一些芥子碱降解效率较高的微生物[10],同时发现,这些微生物对芥子碱的降解可能与一些含铜酶有关。但是,经种属鉴定和安全性评价发现,部分菌种属于条件性致病菌,不能直接应用于生产[11-12]。【拟解决的关键问题】本试验拟通过研究芥子碱、硫酸铜以及培养时间对项目组前期筛选出的一株芥子碱降解优势菌 (Sinapine-degrading Bacteria 1,SDB1)相关酶活力的影响,初步确定SDB1中可能参与芥子碱降解的酶类以及影响酶活的因素,旨在为进一步的应用研究提供理论基础。
1 材料与方法
1.1 试验材料
1.1.1 芥子碱降解菌 芥子碱降解菌(SDB1)是四川省畜牧科学研究院畜禽饲料资源开发项目组前期以芥子碱为唯一碳源,分离培养的一株芥子碱降解优势菌,为肺炎克雷伯氏菌属。
1.1.2 主要试剂与仪器 主要试剂包括:芥子碱硫氰酸盐(购自成都德思特生物技术有限公司,芥子碱含量为80 %);β-葡萄糖苷酶活性测定试剂盒(A031)、脂肪酶活性测定试剂盒(A054)、淀粉酶活性测定试剂盒(C016)、多酚氧化酶活性测定试剂盒(A136)和胆碱酯酶活性测定试剂盒(A023)均购自南京建成生物工程研究所;漆酶活性测试盒(BC1635)购自北京索莱宝科技有限公司。
1.2 芥子碱降解菌的富集培养
使用平板划线法,于优化的脑心浸液琼脂(BHI)培养基中复苏培养SDB1。培养温度27 ℃,培养时间12 h。培养基组成:牛脑200.0 g,牛心浸出汁250.0 g,胰蛋白胨、葡萄糖和酵母提取物适量,NaCl5.0 g,琼脂20.0 g,蒸馏水1000 mL,pH 6.8~7.2。
1.3 试验设计
从SDB1富集培养基中挑取单个菌落接种于SDB1的产酶培养基中。产酶培养基共分为1个对照组和4个处理组,每组4个重复。对照组培养基组成为:NaNO33.0 g;K2HPO41.0 g;MgSO4·7H2O 0.5 g;KCl 0.5 g;FeSO40.01 g;胰蛋白胨和蔗糖适量;水 1000 mL。第1组和第2组是分别在基础培养基中添加0.50和1.00 g/L的芥子碱硫氰酸盐(折算成芥子碱分别为0.40和0.80 g/L),第3组和第4组为分别在基础培养基中添加1.00和3.00 mM/L的五水硫酸铜(表2)。培养基配置好后,调整pH至7.0~7.5,并将每组培养基分装至4个500 mL锥形瓶中(250 mL/瓶)作为4个重复,置于27 ℃振荡培养箱中振荡培养。

表1 试验设计
1.4 菌液收集及处理
取培养第2,4,6,8天的培养液各30 mL菌液,在3000 r/min的条件下离心5~10 min,移除上清液(尽量去除干净),加入PBS液10 mL,使细菌充分重悬于PBS液中。在4 ℃、12 000 r/min的条件下高速离心15 min,收集离心后的上清液,将上清液保存于-20 ℃冰箱中。
1.5 SDB1酶活性鉴定
参照β-葡萄糖苷酶、脂肪酶、淀粉酶、漆酶、多酚氧化酶和乙酰胆碱酯酶活性测试盒操作说明书进行相关酶活力测定。
1.6 统计学分析
结果用平均数±标准差表述,用SPSS统计软件(SPSS v.20.0,IBM Corp.,Armonk,New York,USA)对数据进行方差分析,用t检验对处理组与对照组之间的差异进行显著性检验。
2 结果与分析
2.1 芥子碱和硫酸铜对SDB1淀粉酶活力的影响
由表2可知,SDB1具有一定的淀粉酶活性。与对照组相比,添加0.40 g/L芥子碱,在培养的第2、4、6天,淀粉酶活力均没有显著变化,而在培养到第8天时,淀粉酶的活力却显著高于对照组,是对照组的2.32倍(P<0.05);添加0.80 g/L芥子碱对SDB1淀粉酶活力没有显著影响,但在培养的第4、6、8天,淀粉酶活力均有提高的趋势。此外,添加1.00和3.00 mmol/L的硫酸铜对SDB1淀粉酶活力均没有显著影响,但培养到第4天时2个组的SDB1淀粉酶活力均有增加的趋势。
2.2 芥子碱和硫酸铜对SDB1多酚氧化酶活力的影响
由表3可知,SDB1具有多酚氧化酶活性。尽管与对照组相比,添加0.40 g/L芥子碱对SDB1多酚氧化酶活力没有显著影响,但数值上,多酚氧化酶活力有随培养时间延长而提高的趋势;添加0.80 g/L芥子碱在培养的第2和4天,多酚氧化酶活性极显著高于对照组,分别比对照组增加了75.70 %(P<0.01)和197.03 %(P<0.01)。添加1.00和3.00 mmol/L的硫酸铜对SDB1多酚氧化酶活力均没有显著影响。
2.3 芥子碱和硫酸铜对SDB1β-葡萄糖苷酶活力的影响
由表4可知,SDB1几乎不具有β-葡萄糖苷酶活性。与对照组相比,添加0.40和0.80 g/L的芥子碱对SDB1β-葡萄糖苷酶活力均没有显著影响。添加1.00和 3.00 mmol/L的硫酸铜对SDB1中β-葡萄糖苷酶活力均没有影响。
19.紧扣重要时间节点开展集中宣传。鼓励探索重要节点开展法治宣传的新模式,充分利用“4·26”知识产权日、宪法宣传周等时间点,选取民营企业关注的法律法规或法律热点问题,集中开展大型的普法宣传活动,借助重要节点影响力进一步提升普法效果,提高工作覆盖面,传播法律知识,弘扬法治精神。

表2 芥子碱和硫酸铜对SDB1淀粉酶活力的影响
注:同行肩注无*表示不显著(P>0.05),有*表示与对照组相比差异显著(P<0.05),有**表示与对照组相比差异极显著(P<0.01),下同。
Notes:In the same row, values with no asterisk superscripts mean no significant difference (P>0.05)between the control group and the treatment groups, while with one asterisk(*) superscripts mean significant difference (P<0.05) between the control group and the treatment groups, and with two asterisk (**) superscripts mean very significant difference (P<0.01) between the control group and the treatment groups. The same as below.

表3 芥子碱和硫酸铜对SDB1多酚氧化酶活力的影响
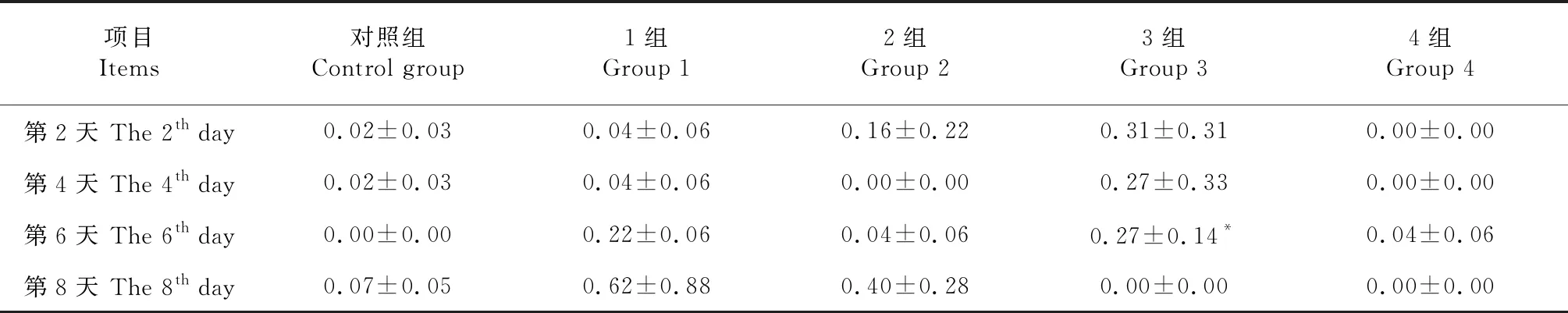
表4 芥子碱和硫酸铜对SDB1β-葡萄糖苷酶活力的影响
2.4 芥子碱和硫酸铜对SDB1漆酶活力的影响
由表5可知,SDB1具有较高的漆酶活性,且漆酶活性随培养时间延长而增加。与对照组相比,添加0.40和0.80 g/L的芥子碱均可以提高SDB1漆酶活力,其中添加0.40 g/L的芥子碱,在培养第4天,SDB1漆酶活力极显著提高,是对照组的6.25倍(P<0.01);添加0.80 g/L的芥子碱,漆酶活力在培养第4和6天均极显著提高,在培养的第8天也显著提高,分别为对照组的5.92倍(P<0.01)、3.03倍(P<0.01)和5.70倍(P<0.05)。添加1.00和3.00 mmol/L的硫酸铜对SDB1漆酶活力均没有显著影响。
2.5 芥子碱和硫酸铜对SDB1乙酰胆碱酯酶活力的影响
由表6可知,SDB1具有一定的乙酰胆碱酯酶活性。与对照组相比,添加0.40 g/L的芥子碱对SDB1乙酰胆碱酯酶活力没有影响;添加0.80 g/L的芥子碱,乙酰胆碱酯酶的活性在培养第6天时显著降低,其余培养时间没有显著变化。添加1.00和3.00 mmol/L的硫酸铜对SDB1乙酰胆碱酯酶活力没有显著影响。
2.6 芥子碱和硫酸铜对SDB1脂肪酶活力的影响
由表7可知,SDB1具有一定的脂肪酶活性。与对照组相比,添加0.40和0.80 g/L的芥子碱以及1.00和3.00 mmol/L的硫酸铜对SDB1脂肪酶活力均没有显著影响。
3 讨 论
芥子碱是芥子酸与胆碱形成的酯类化合物,其化学名称为4-羟基-3,5二甲氧基苯丙烯胆碱酯。从分子结构看,芥子碱的降解有3个潜在的活性位点:酯基、苯环上的酚羟基和碳碳双键。酯键可以直接水解为芥子酸和胆碱;酚羟基可能发生氧化反应形成醛酮或者醌类物质,双键也可能发生环氧化反应[13]。近年来,芥子碱的微生物降解研究主要集中在真菌上:肖翼[14]用米曲霉固态发酵法有效脱除了菜籽粕中的芥子碱和多酚。Lacki[15-16]用白腐真菌有效降解了菜粕芥子碱的含量。Rozan[17]采用少孢根霉也成功降解了菜籽粕30 %的酚类物质。周浩宇[8]用酿酒酵母、纳豆芽孢杆菌及枯草芽孢杆菌B4混合发酵菜籽饼粕,使菜籽粕中的芥子碱降解率达到80.06 %。而钮琰星[13]指出酿酒酵母无论是液态发酵还是固态发酵都对菜籽粕芥子碱含量没有显著影响。本试验采用的SDB1为项目组前期分离培养的一株芥子碱降解优势菌,与项目组前期报道的芥子碱降解菌均为肺炎克雷伯氏菌属[10],具有显著的芥子碱降解效果。与前人报道不同的是,SDB1属于细菌类微生物。与真菌相比,细菌基因简单,新陈代谢旺盛,培养条件要求不高,能够在较短时间内进行大量繁殖,具有潜在的优势。
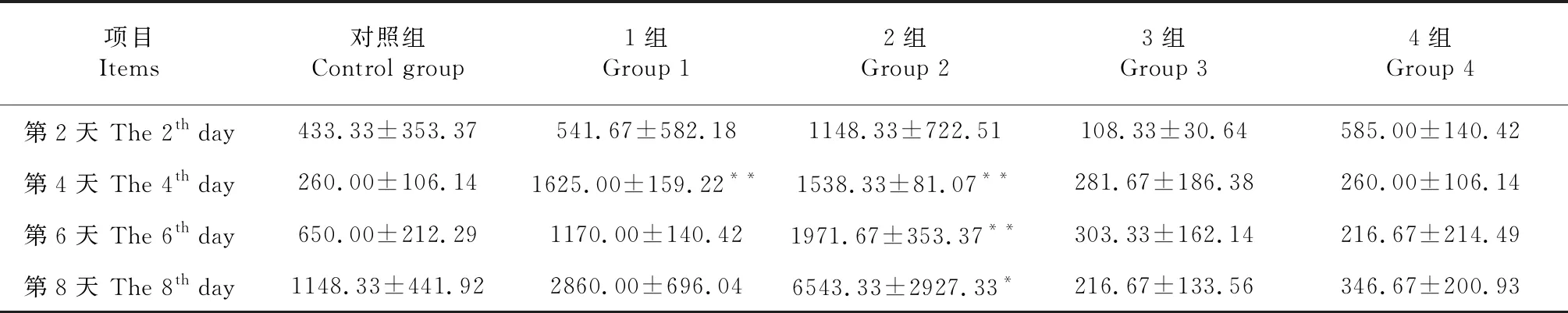
表5 芥子碱和硫酸铜对SDB1漆酶活力的影响

表6 芥子碱和硫酸铜对SDB1乙酰胆碱酯酶活力的影响

表7 芥子碱和硫酸铜对SDB1脂肪酶活力的影响
微生物降解芥子碱的实质是其分泌的生物酶对芥子碱的降解。目前研究较多的酶包括多酚氧化酶、漆酶、酪氨酸酶、单宁酸酶、阿魏酸酯酶、β-葡萄糖苷酶和脂肪酶等[18-20]。多酚氧化酶是自然界中分布极广的一类氧化还原酶,是目前已知的可以有效降解芥子碱的主要酶系。Lacki指出白腐真菌分泌的多酚氧化酶可以有效降解芥子碱,其降解效果与温度、pH和氧气浓度有关。用相同的酶降解芥子酸也会出现类似的结果[15-16],说明多酚氧化酶对芥子碱的作用位点可能在芥子碱中芥子酸的部分。本试验对照组多酚氧化酶活力在数值上随培养时间延长而提高,说明SDB1具有多酚氧化酶活性。添加0.40 g/L芥子碱的后,尽管多酚氧化酶活力与对照组相比没有显著差异,但是在数值上提高的趋势明显,到培养的第8天时,已经达到对照组的2.11倍。而当芥子碱的添加量达到0.80 g/L时,多酚氧化酶的活性在培养的第4和6天极显著高于对照组(P<0.01),说明芥子碱作为底物充分激活了SDB1的多酚氧化酶。作为一种铜依赖性蛋白,多酚氧化酶活性受铜离子影响较大,但本试验添加1.00和3.00 mmol/L的硫酸铜,对SDB1多酚氧化酶活性却没有显著影响,原因有待进一步研究。
微生物来源的多酚氧化酶主要是漆酶和酪氨酸酶[21]。漆酶是多酚氧化酶中作用底物最广的一类。Hu研究了固态发酵过程中的漆酶活性和芥子碱的含量,发现漆酶的产生对芥子碱含量的降低具有显著作用[22]。微生物漆酶按来源分为真菌漆酶和细菌漆酶[23]。真菌漆酶广泛存在于多孔菌、脉孢菌、曲霉菌等真菌属中, 其中担子菌中的白腐菌漆酶研究最多[24]。钮琰星[13]的研究指出来源于云芝菌的漆酶对菜籽粕芥子碱具有显著的酶解作用,且漆酶的反应活性位点是苯环的酚羟基。细菌漆酶研究不如真菌漆酶深入,目前的报道以芽孢杆菌属为主,生脂固氮螺菌、链霉菌、大肠杆菌等细菌中也发现过漆酶活性[25]。与真菌漆酶相比,细菌漆酶可能具有更多的优点, 如它具有Cu2+抗性[26]、不需糖基化、热稳定性好[27]、最适pH值范围广[28]等。因此, 细菌漆酶的应用前景更为广阔。本试验采用的SDB1就属于细菌类微生物,试验结果显示:对照组漆酶活力在数值上随培养时间延长而提高,第8天的漆酶活力为第2天的2.65倍,说明SDB1具有较强的漆酶活性。该结果也为细菌漆酶验证了一位新成员。添加0.40或0.80 g/L芥子碱后,SDB1的漆酶活力在培养的第4天到第8天内显著或极显著高于与对照组(P<0.01),说明芥子碱作为底物充分激活了SDB1的漆酶。与多酚氧化酶相同,漆酶是一类典型含铜糖蛋白,铜离子是其催化反应的活性中心,因此其活性还会受铜离子含量直接影响。范晶晶[29]研究指出在培养基中添加0.50~4.00 mmol/L硫酸铜,均能提高SYBC starX菌(为Kocuria菌属)的漆酶活性,其中最适Cu2+浓度为3.00 mmol/L。但是本试验在培养基中添加1.00和3.00 mmol/L的硫酸铜后,SDB1漆酶活力却没有增加。目前研究较多的细菌漆酶是CotA蛋白,它来源于芽孢杆菌。汪春蕾证实CotA存在于芽孢杆菌的芽孢壁[30],是一种铜依赖性蛋白[31]。Matthias将CotA蛋白在大肠杆菌中异源表达发现,除了胞内铜离子浓度外,适宜的铜伴侣也是影响铜离子嵌入CotA蛋白分子并影响CotA活性的重要因素[32]。本试验中硫酸铜没有增加漆酶和多酚氧化酶活性是否也与没有响应的铜伴侣有关,目前还不清楚,需要进一步的研究。尽管芽孢杆菌漆酶是目前细菌漆酶的典型代表,但由于漆酶一般存在于芽孢杆菌的芽孢壁中,提取成本较高,在生产中应用受限,因此筛选具有胞外产漆酶能力强的细菌,对于细菌漆酶的应用具有十分重要的意义。本试验的SDB1漆酶就是一种胞外细菌漆酶,在推广应用上具有很大优势。
何江[20]的研究结果表明,β-葡萄糖苷酶和脂肪酶也参与菜籽粕芥子碱的降解,只是降解效果不如漆酶和多酚氧化酶。周浩宇[19]指出酿酒酵母中的β-葡萄糖苷酶可以降解17.65 %的菜籽粕芥子碱。本试验也研究了芥子碱和硫酸铜对SDB1的β-葡萄糖苷酶和脂肪酶的活性。结果对照组SDB1不具有β-葡萄糖苷酶活性,具有一定的脂肪酶活性。添加芥子碱和硫酸铜没有显著改变SDB1的β-葡萄糖苷酶和脂肪酶活性。由于现有大多数研究都是用菜籽粕替代芥子碱,仅仅是通过测定发酵前后菜籽粕芥子碱的含量变化来推测酶与芥子碱的关系,并没有考虑菜籽粕复杂的物质组成可能对酶活性和芥子碱含量产生的影响;而本试验是直接采用的芥子碱硫氰酸盐(含量达80 %以上),这可能是导致本试验结果与现有研究出现差异的原因之一。关于芥子碱与β-葡萄糖苷酶以及脂肪酶的关系目前报道极少,以上推测有待进一步验证。
项目组前期的抑菌圈定性试验表明研究[10]表明:与SDB1同种属的YD-1和YD-2均具有淀粉酶活性。本试验也进一步考察了SDB1的淀粉酶活性,并研究了芥子碱和硫酸铜对淀粉酶活性的影响。结果SDB1具有一定的淀粉酶活性。添加0.40 g/L芥子碱,SDB1淀粉酶的活力在培养到第8天时显著高于对照组;添加0.80 g/L芥子碱也有提高淀粉酶活力的趋势。添加硫酸铜对SDB1淀粉酶活力没有显著影响。目前还未见其它关于芥子碱与淀粉酶活力关系的研究报道。结合本试验脂肪酶和淀粉酶的结果可以设想,当SDB1用于消减十字花科类饲料资源中的芥子碱时,还有可能在一定程度上改善碳水化合物和脂类养分的消化利用率,从而多方面改善饲料品质。
4 结 论
(1) 本试验条件下,SDB1具有一定的淀粉酶、多酚氧化酶、漆酶、乙酰胆碱酯酶和脂肪酶的活性,不具有β-葡萄糖苷酶的活性。
(2)本试验条件下,培养基中添加芥子碱,可以在培养的4~8 d内不同程度的提高SDB1淀粉酶、多酚氧化酶、漆酶活性;芥子碱添加量越高,多酚氧化酶和漆酶活力越高;添加芥子碱对SDB1的β-葡萄糖苷酶、乙酰胆碱酯酶、脂肪酶活性没有显著影响。
(3) 本试验条件下,培养基中添加硫酸铜对SDB1的淀粉酶、多酚氧化酶、漆酶、β-葡萄糖苷酶、乙酰胆碱酯酶和脂肪酶活性均没有显著影响。
猜你喜欢
农产品加工(2022年15期)2022-09-26
中华胰腺病杂志(2022年4期)2022-08-23
现代农业科技(2022年12期)2022-07-02
水产科学(2021年4期)2021-07-21
渔业致富指南(2020年19期)2020-11-10
求知导刊(2019年15期)2019-08-30
故事会(2019年1期)2019-01-11
广西科技大学学报(2018年2期)2018-09-10
雪莲(2016年2期)2016-05-26
东方艺术·国画(2015年3期)2015-08-20
