
彩叶杨树叶片中花色素苷的光保护作用
2020-06-22 08:28朱秀红杨金橘温道远韩晓雪茹广欣
西南农业学报 2020年5期
朱秀红,杨金橘,温道远,韩晓雪,茹广欣
(河南农业大学林学院,河南 郑州 450000)
【研究意义】中华红叶杨是首个由黑杨2025雄株芽变产生的彩叶杨树品种,推向市场以来彩叶杨树品种的丰富度与观赏度不断提高,以其色彩的缤纷斑斓,极强的适应能力以及较快的生长周期广受喜爱,而花色素苷作为彩叶植物成色的关键因素,研究不同品种彩叶杨树花色苷合成机制及其在不同环境胁迫下对叶片保护作用的调控机理,对促进我国进一步丰富以花色素苷呈色为基础的彩叶树品种具有重要意义。【前人研究进展】花色素苷不止可以为植物提供丰富多彩的颜色,已有研究还证明作为多酚类物质的花色素苷能够以其特殊结构减弱光照胁迫引起的过量ROS产生,对叶片起到一定的光保护作用[1-2]。【本研究切入点】通过研究光照胁迫期间及恢复后5个彩叶杨树品种的花色素苷含量、叶绿素含量、类胡萝卜素含量、叶黄素循环、最大光化学效率(Fv/Fm)及相关酶活性和非酶抗氧化剂活性等指标的差异,以探究叶色变化对于植物叶片光能的吸收与利用造成的影响,证明高光胁迫引起的花色素苷含量与积累与相关抗氧化酶活性,叶内ROS以及非酶抗氧化剂活性(DPPH自由基清除能力)的相互关系。【拟解决的关键问题】深入研究过剩光能诱导花色素苷的合成机制,以期为彩叶杨树的广泛栽培和应用提供进一步理论指导。
1 材料与方法
1.1 材料与设计
在新密试验田中,选取长势良好的全红杨、中红杨、金红杨、红霞杨与翡翠杨的一年生苗木同侧枝剪下带回进行扦插培养(每个品种每株重复扦插3盆),幼苗生长至25 cm(包括顶芽,主枝生3~4片叶 )放置于温室内,温度为15~25 ℃,每天遮光10 h继续培养,确保可取新鲜样品进行重复试验。
试验以常温常光(CK)为对照(温度为 25/15 ℃,光照强度为 200 μmol·m-2·s-1),用高光胁迫(温度为 25/15 ℃,光照强度为500 μmol·m-2·s-1)连续处理3 d,每天进行取样测定,3 d后将受胁迫后样本材料放置于智能温室内进行适当遮阴处理恢复光照,在恢复的2 d进行取样测定。
1.2 试验方法
采用张宪政提出的丙酮乙醇萃取法[3]进行叶绿素与类胡萝卜素含量测定。采用何奕昆[4],于晓南[5]等的盐酸甲醇分光光度计方法,进行花色素苷的测定。分别取5种彩叶杨树顶芽向下第2片叶进行叶黄素各组分含量的测定。采用THAYER等[6]的方法用高效液相色谱法测定。叶黄素循环的脱环氧化程度用(A+Z)/(V+A+Z)表示。用便携式脉冲调制荧光仪FMS22测定5种彩叶杨树叶片的初始叶绿素荧光(Fo)及暗适应下的最大叶绿素荧光(Fm),用Fv/Fm表示PSII的最大光化学效率值,其中Fv由(Fm-Fo)计算所得[7]。用过氧化氢紫外分光光度法[8]测定过氧化氢酶(CAT)的活性;用愈创木酚法测定过氧化物酶(POD)活性[8];用 NBT(氮蓝四唑)光化学还原法测定超氧化物歧化酶(SOD)活性;采用赵世杰等改进紫外吸收法测定抗坏血酸还原酶(APX)活性[9];采用De Gara L 等提出的磷酸缓冲法进行DHAR活性测定[10]。H2O2含量采用刘俊等(2000)的离心标准曲线方法[11]。超氧离子产生速率采用王爱国等的方法[12]。DPPH测定采用张宏梅等改进的紫外分光光度法[13]。
1.3 数据分析
采用Origin9.0与Excel2016软件对实验数据进行统计与分析,并用单因素方差分析来检验样本间的差异显著性(置信区间为95 %)。本文中的折线图以及柱状图均为平均值,标准差为误差线。
2 结果与分析
2.1 Fv/Fm和ROS在高光胁迫下的变化
在胁迫开始的第1天,5个彩叶杨树品种的最大光化学效率(Fv/Fm)差异并不显著,在0.65~0.75范围内(图1)。随着高光胁迫处理的时间(天数)增长,5个品种的Fv/Fm均开始下降,即在叶片内都发生了光抑制现象。且根据图1可知,翡翠杨(绿叶品种)的Fv/Fm值下降最快,全红杨的Fv/Fm值变化幅度最小,中红杨、金红杨与红霞杨的下降幅度介于两者之间。在胁迫开始的第3天,全红杨的Fv/Fm值最高,是翡翠杨的1.63倍。进行光照恢复后,5个品种的Fv/Fm逐渐上升,光照恢复第1天,全红杨的最大光化学效率已恢复至胁迫初始水平,其余4个品种均上升至略低于初始水平。光照恢复后的第2天,全红杨和中红杨的Fv/Fm值上升到略高于原始水平,翡翠杨的Fv/Fm值恢复到胁迫初始值的85 %,金红杨和红霞杨恢复至与初始水平相当。表明5个品种中,全红杨对高光胁迫有较强的抵御作用,能更好的对光照胁迫做出应答。而翡翠杨的最大光化学效率(Fv/Fm)随光照强度变化显著,在高光胁迫时叶片内遭受了更强烈的光系统破坏。
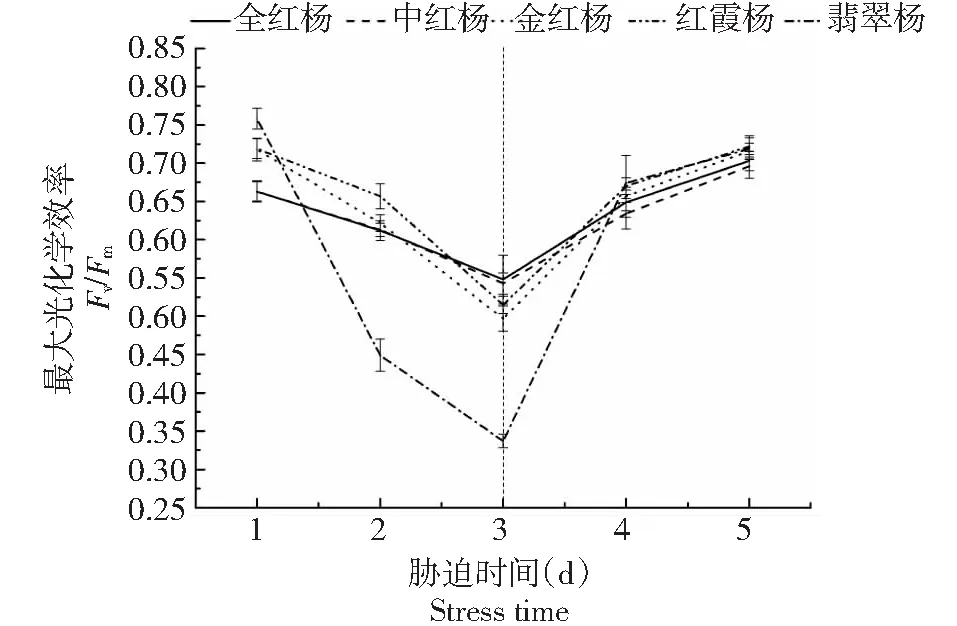
图1 高光胁迫下5个彩叶杨树品种的最大光化学效率变化Fig.1 Changes in Fv/Fm in leaves of five colorful poplar varieties in response to high-light stress
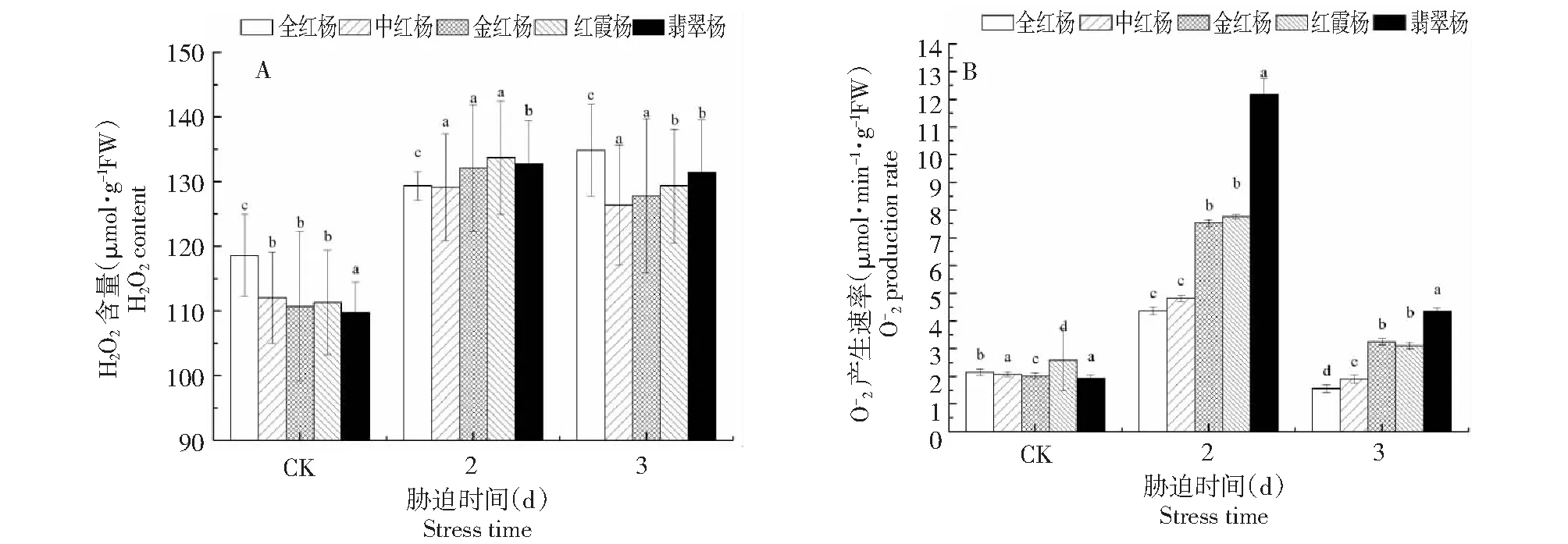
图2 高光胁迫下5个彩叶杨树品种的H2O2含量(A)和产生速率(B)变化Fig.2 Changes in H2O2 content and production rate in leaves of five colorful poplar varieties in response to high-light stress
2.2 色素含量在高光胁迫下的变化
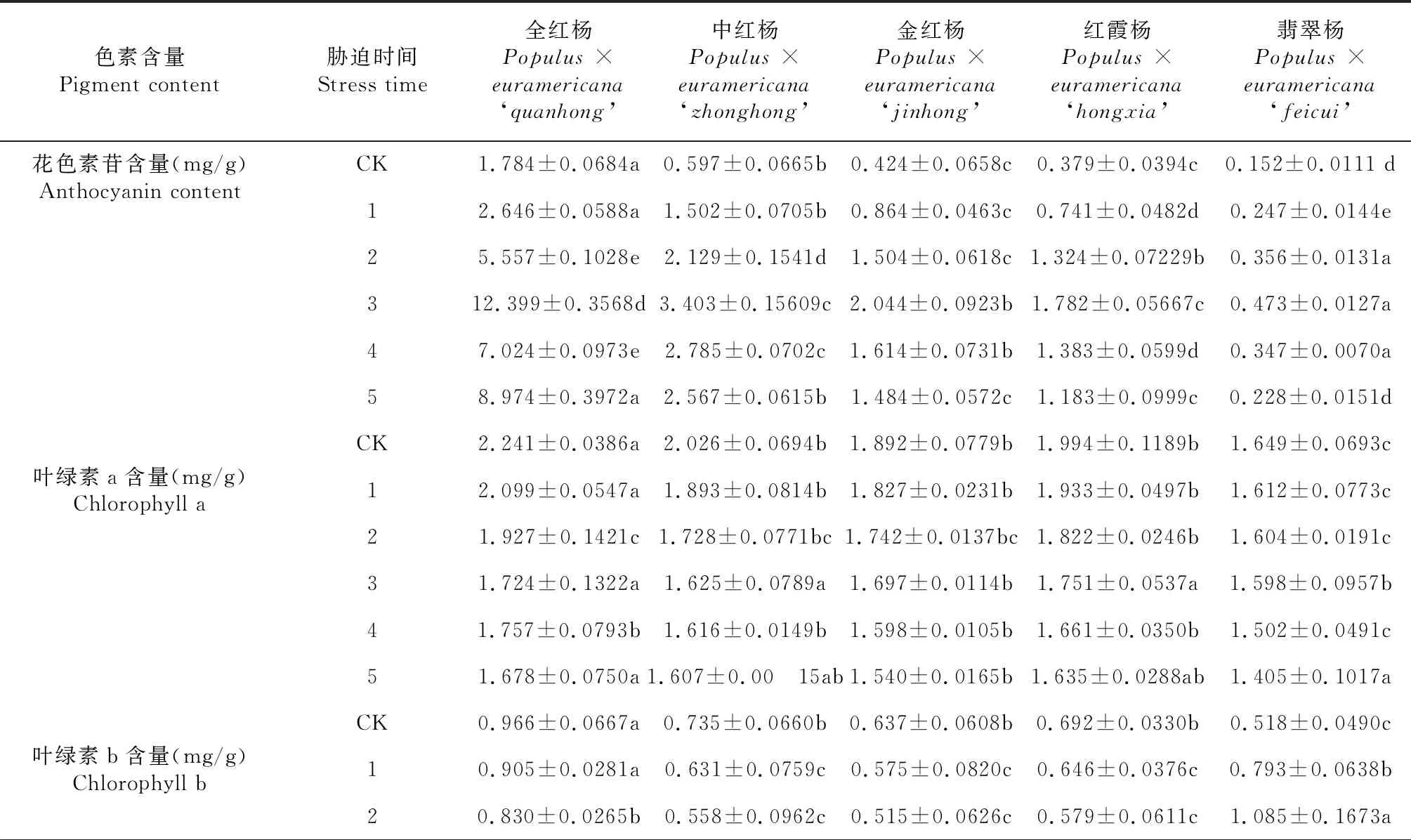
表1 高光胁迫下5个品种的彩叶杨树的花色素苷、叶绿素及类胡萝卜素含量变化
续表1 Continued table 1

色素含量Pigment content胁迫时间Stress time全红杨Populus ×euramericana ‘quanhong’中红杨Populus ×euramericana ‘zhonghong’金红杨Populus ×euramericana ‘jinhong’红霞杨Populus ×euramericana ‘hongxia’翡翠杨Populus ×euramericana ‘feicui’30.743±0.0903b0.424±0.1377c0.424±0.0691c0.586±0.0495bc1.374±0.0747a40.657±0.0554c0.341±0.0563b0.406±0.0852a0.341±0.1202c1.012±0.0694b50.723±0.0406a0.328±0.0448bc0.433±0.1017b0.272±0.1039c0.443±0.0382bCK3.207±0.0877a2.761±0.1353b2.529±0.1153 b2.686±0.1518b2.167±0.1181 c叶绿素含量(mg/g)Chlorophyll content13.004±0.0266a2.524±0.0548bc2.402±0.0798c2.579±0.0803b2.405±0.1013c22.757±0.1633a2.286±0.0694b2.257±0.0511b2.401±0.0849a2.689±0.1545a32.467±0.1191b2.049±0.0590c2.121±0.0799c2.337±0.1026b2.972±0.1474a42.414±0.1064a1.957±0.0710b2.004±0.0785b2.002±0.0972c2.514±0.1167b52.401±0.1149a1.935±0.0502b1.973±0.0894b1.907±0.0763b1.848±0.1339bCK0.712±0.0180a0.663±0.0502ab0.595±0.0407c0.612±0.0271bc0.501±0.0219d类胡萝卜素含量(mg/g)Carotenoid content10.792±0.0376a0.707±0.0132b0.622±0.0131c0.649±0.0108c0.556±0.0554d20.813±0.0164a0.725±0.0066b0.701±0.0101b0.727±0.0098b0.611±0.0816c30.845±0.0286ab0.796±0.0115ab0.735±0.0076c0.784±0.0061bc0.665±0.0584d40.701±0.0197c0.671±0.0113ab0.554±0.0104c0.635±0.0079b0.581±0.0562c50.619±0.0157a0.589±0.0106ab0.546±0.0085b0.565±0.0082ab0.477±0.0699c
在对照组中,全红杨的花色素苷含量是翡翠杨的11.72倍(表1),进行光照胁迫后,5个品种的花色素苷含量均显著增加,但翡翠杨的花色素苷含量变化幅度的相较于其余4个品种小,胁迫后的第3天,全红杨的花色素苷含量增加至翡翠杨的26.21倍。恢复2 d后,翡翠杨的花色素苷含量降至与对照水平相当,中红杨的花色素苷含量下降至对照水平的4.30倍,金红杨与红霞杨恢复后下降至对照水平的3倍左右,全红杨维持了胁迫诱导后积累的大部分花色素苷。
5个品种的叶绿素a与叶绿素b含量在对照组中差异不显著,进行胁迫的第1天,全红杨叶绿素a含量是翡翠杨的1.3倍,中红杨,金红杨与红霞杨的叶绿素a含量维持在1.82~1.94 mg/g范围内,差异并不显著。进行光照胁迫后,5个品种的叶绿素a含量均呈不明显的下降趋势。进行恢复的第2天,翡翠杨的叶绿素a含量下降至1.40 mg/g,其余4个品种均维持在1.60 mg/g左右。全红杨,中红杨,金红杨与红霞杨的叶绿素b含量在胁迫后同样呈不显著的下降趋势,而值得注意的是翡翠杨的叶绿素b含量在高光胁迫过程中却出现缓慢上升,在胁迫的第3天,叶绿素b含量上升至对照水平的2.65倍;恢复光照后,翡翠杨的叶绿素b含量开始下降,恢复的第2天下降至对照的85.50 %。叶绿素含量与组成成分的比值是衡量植物光捕获能力的标准之一,虽然5个彩叶杨树品种的花色素苷含量差异显著,但光捕获能力在遭受高光胁迫及恢复后并未出现显著差异。

图3 高光胁迫下5个彩叶杨树品种的叶黄素循环库(C)和脱环氧化程度(D)Fig.2 Changes in xanthophyll pool size(V+A+Z) and de-epoxidation state[(A+Z)/(V+A+Z)]in leaves of five colorful poplar varieties in response to high-light stress
5个品种的类胡萝卜素含量在光照恢复后与对照水平基本一致,但高光胁迫期间使5个品种的类胡萝卜素出现了不同程度的下降,全红杨在胁迫后与恢复后都具有较高的类胡萝卜素含量。
2.3 叶黄素变化
叶黄素循环可以表现植物的热耗散效率,对热能的耗散使植物具有一定的光保护作用。从图3可以看出,高光胁迫对全红杨的叶黄素库容量(V+A+Z)没有显著影响(C),其余4个品种在光照胁迫下呈上升趋势,其中翡翠杨的叶黄素库容量(V+A+Z)上升幅度最大。光照恢复后5个品种的叶黄素库容量(V+A+Z)均下降至对照水平。从图3(D)可以知道,高光胁迫对5个品种的叶黄素脱环氧化程度(A+Z)/(V+A+Z)影响较为显著,进行胁迫3 d后,翡翠杨的(A+Z)/(V+A+Z)上升至对照的1.67倍,全红杨和中红杨的(A+Z)/(V+A+Z)的变化幅度略小,增长1.45倍左右,金红杨和红霞杨的增幅均为1.5倍左右。即在高光胁迫下,5个品种的叶黄素脱环氧化增长程度类似,进行光照恢复后,5个品种均恢复至略高于对照水平,其中翡翠杨的叶黄素脱环氧化程度在光照恢复后保持在对照值的1.33倍。综合对比可以知道,虽然翡翠杨的(A+Z)/(V+A+Z)值高于其余4个品种,但全红杨在具有较高的叶黄素脱环氧化程度的同时,其叶黄素库容量(V+A+Z)高于翡翠杨,因此全红杨具有较高的热耗散效率。
2.4 抗氧化剂和抗氧化酶活性变化
从图4可以看出,全红杨的DPPH清除能力在胁迫前最高,随着光照胁迫时间的延长显著增加,恢复阶段维持平缓下降。中红杨、金红杨和红霞杨的DPPH清除能力依次下降,在胁迫下的增加幅度也依次减小,恢复2 d后能维持最初水平。翡翠杨的DPPH清除能力相比于其他4个品种较低,且在整个胁迫阶段和恢复阶段都没有显著变化。说明,全红杨,中红杨等红叶品种具有较高的非酶抗氧化剂活力,对其光保护能力有较大贡献,而翡翠杨的非酶抗氧化剂活力较低。
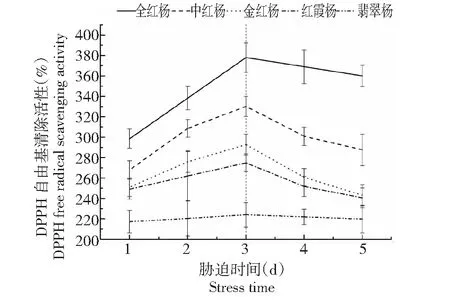
图4 高光胁迫下5个彩叶杨树品种的非酶抗氧化剂DPPH清除能力Fig.4 Changes in radical scavenging activities in leaves of five colorful poplar varieties in response to high-light stress
由图4与表1的对比可得,随着花色素苷含量的增加,5个品种的DPPH活性也不断上升。表明在高光胁迫与光照恢复阶段,基于DPPH的自由基清除能力与花色素苷含量密切相关,花色素苷存在下,叶片的光保护能力主要表现为对非酶抗氧化剂的促进作用。
在对照组中(胁迫前),5个品种的SOD、POD、CAT和APX 4种抗氧化酶活性差异并不显著(图5-E,F,G,I),脱氢抗坏血酸还原酶(DHAR)的差异较为明显(图4-H)。高光胁迫使5个品种的彩叶杨树叶片内的5种抗氧化酶活性有不同程度的提高。全红杨与翡翠杨的SOD,POD活性相当,在进行光照胁迫后,两者的酶活性增加幅度也相同,但光照恢复以后,翡翠杨的POD维持在对照水平的171.24 %,而全红杨下降至对照的76 %,其余3个品种在光照恢复后均下降至对照水平。
翡翠杨的过氧化氢酶(CAT)和脱氢抗坏血酸还原酶(DHAR)在高光胁迫下的增加幅度基本相同,其增幅与含量均显著高于全红杨。中红杨,金红杨和红霞杨CAT和DHAR活性介于2个品种之间,且在光照恢复后都恢复至对照水平。值得注意的是,与其余4个品种相比,翡翠杨的抗坏血酸过氧化物酶(APX)在胁迫后显著上升,且在恢复光照强度后仍维持了较高APX含量,而其余4个品种的APX活性随着光照强度的减弱下降至对照水平。
综合图4~5可以知道,全红杨红叶品种对高光胁迫的良好应答机制主要归功于较高的非酶抗氧化剂活力(DPPH自由基清除能力),而抗氧化酶并未对其光照保护能力表现出突出贡献。翡翠杨(绿叶品种)对高光胁迫的应答机制主要依靠抗氧化酶含量及活性的增强。
3 结论与讨论
3.1 高光胁迫诱导产生的过剩光能
与对照组相比,维持相同温度条件下,光照强度增强导致5个品种彩叶杨树的Fv/Fm值明显下降,表明叶片的最大光化学效率下降,光系统的反应中心遭到损伤[14-15]。
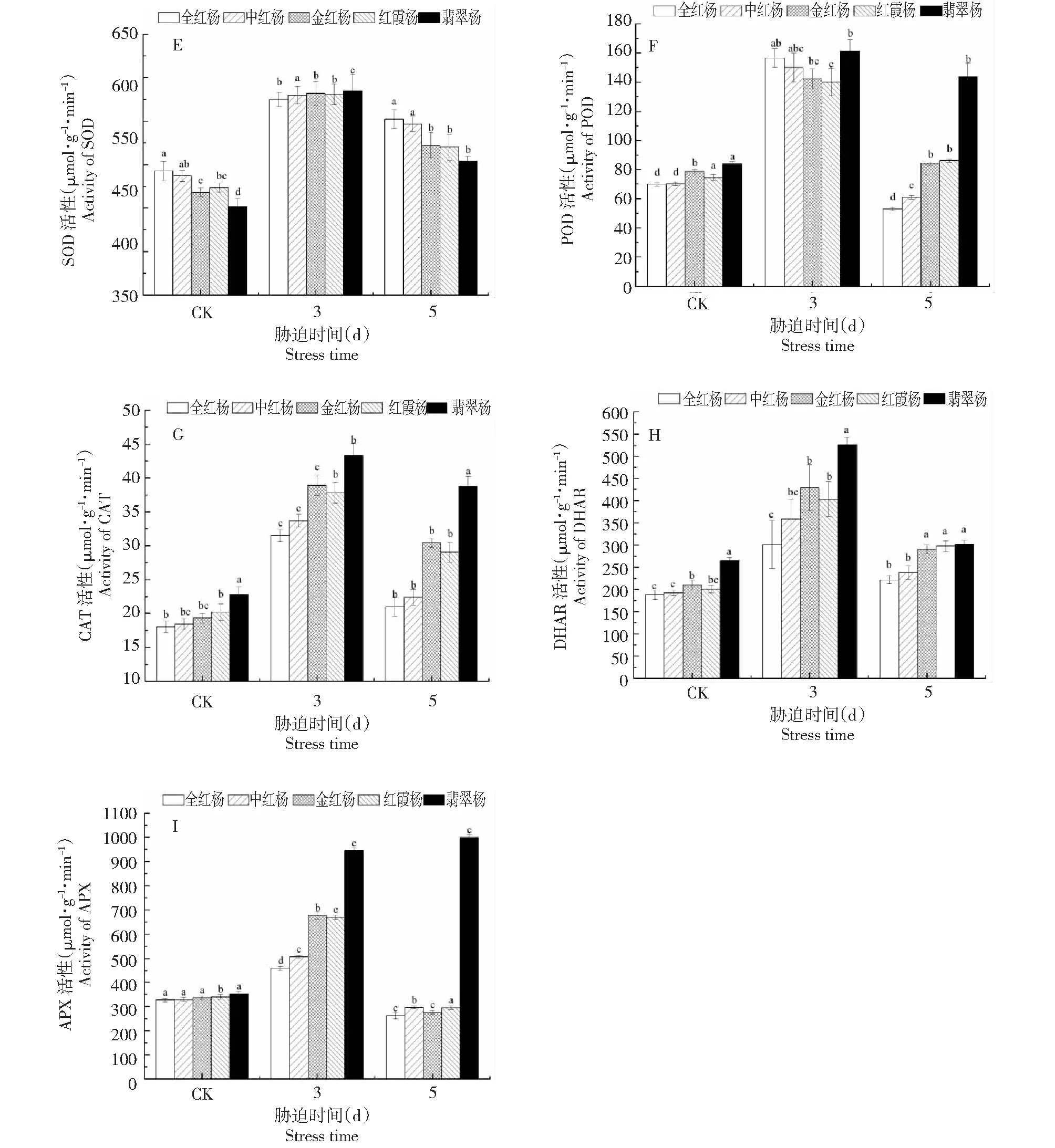
图5 高光胁迫下5个彩叶杨树品种的抗氧化酶活性变化Fig.5 Changes in antioxidant enzyme activities in leaves of five colorful poplar varieties in response to high-light stress
3.2 高光胁迫下花色素苷的产生与积累
3.3 花色素苷在彩叶杨树中的光保护作用
Chalker-Scott L在花色素苷对环境胁迫的响应一文中已讨论,出现环境胁迫时(例如本文中利用高光胁迫),植物在各组织中会积累一定的花色素苷[25],这为花色素苷与叶片光保护作用之间存在密切关系提供了证据。且后有研究已报道,遭受低温及光照等非生物胁迫后均可以导致花色素苷的进一步合成,同时增加积累量[26-27]。本文中,光照胁迫下花色素苷的大量合成与积累以及各抗氧化酶活性和非酶抗氧化剂活性的显著变化,均证明高光胁迫下花色素苷含量越高的品种对叶片的光保护能力越强。同时,进行光照胁迫后,5个品种的叶绿素含量及组成比例变化未出现明显变动,即它们的光捕获能力在遭受高光胁迫及恢复后并未出现显著差异。侧面说明,叶片此时出现的光保护作用并非来自光捕获能力的贡献,而主要源自于花色素苷的大量合成与积累,这为彩叶杨树中含有花色素苷含量更多的品种具有更强的光保护能力提供了进一步证明。
猜你喜欢
中老年保健(2022年3期)2022-08-24
现代园艺(2021年16期)2021-12-06
华人时刊(2021年15期)2021-11-27
江西农业(2021年2期)2021-03-19
今日农业(2020年15期)2020-09-25
流行色(2020年5期)2020-06-29
NBA特刊(2018年10期)2018-06-02
NBA特刊(2018年10期)2018-06-02
NBA特刊(2018年10期)2018-06-02
唐山文学(2016年11期)2016-03-20
