
Temporal and spatial variations of bacterial community compositions in two estuaries of Chaohu Lake*
2020-06-08 05:33ZHANGLeiLILiLIUMengnaHUYaoJIANGJiahu
ZHANG Lei , , LI Li , LIU Mengna , HU Yao , JIANG Jiahu
1 State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008
2 School of Civil Engineering and Architecture, Chuzhou University, Chuzhou 239000, China
Received Apr. 19, 2019; accepted in principle Aug. 5, 2019; accepted for publication Sep. 6, 2019 © Chinese Society for Oceanology and Limnology, Science Press and Springer-Verlag GmbH Germany, part of Springer Nature 2020
A bstract The distinctive estuary hydrodynamics and nutrient input make the estuary ecosystem play a key role in lake ecosystems. The Nanfei River and Zhaohe River are two main inlets of Chaohu Lake, Anhui, East China. We selected estuaries of the two rivers as representative areas to study temporal and spatial changes of bacterial communities. In August (summer) and November (autumn) 2016 and February (winter) and May (spring) 2017, 16 water and sediment samples were collected from the estuaries. Physicochemical characteristics indicate signifi cant difference erences in the nutritional status and eutrophication index of the estuaries due mainly to organic input. Examination of the number of operational taxonomic units, the diversity index, the community composition, and redundancy analysis revealed the following. First, the existence of varying degrees of seasonal difference erences in the distribution of almost all bacteria. In addition, the species diversity in the sediment samples was higher than that in the water samples, and the dominant species difference ered also among these samples. Second, a large number of unknown genera were detected, especially in the sediment samples, such as unclassifi ed Xanthomonadales incertae sedis, unclassifi ed Anaerolineaceae, and unclassifi ed Alcaligenaceae. Last, TP, TN, and TOC were the main infl uential factors that afference ected the bacterial community structure.
K eyword: estuary; water and sediment; bacterial diversity; bacterial community structure
1 INTRODUCTION
Pollutants in estuarine waters originate mainly from local river inputs due to human activities. When a river enters a lake, the hydrodynamic efference ect weakens, and pollutants in the lake may combine with particles through adsorption and sedimentation (Eggleton and Thomas, 2004), causing difference erence in nutrient distribution between sediment and water of the lake. As an important component of an ecosystem, lakes are rich in microbial resources, of which heterotrophic prokaryotes play a crucial role in the degradation and redistribution of chemical pollutants (Fernández-Luqueño et al., 2011; Liu et al., 2011). Studies on the efference ects of human activities and environmental conditions on bacterial community structures and organic mineralization have been carried out in lake ecosystems, including eutrophic shallow lakes (Tang et al., 2015), large inland saltwater lakes (Zhang et al., 2016), oligotrophic open oceans (Fuhrman et al., 1993), and cold spring sediments (Mills et al., 2004). These studies helped to uncover the link between bacterial community structure and lake nutrient status and to improve the understanding of bacterial diversity in difference erent environments (Cao et al., 2014).
Chaohu Lake (117°17′-117°52′E, 31°25′-31°43′N) is one of the fi ve largest freshwater lakes of China, total surface area 780 km2and average depth 2.7 m (Deng et al., 2007). Chaohu Lake plays many roles in water supply, fl oods control, ecological balance, and economic development (Meng and Ma, 2015). However, as a shallow lake, the lake is very susceptible to human activities. In recent decades, with the quick economic development of surrounding cities and towns, wastewaters from industry and household polluted the lake via rivers, introducing a large volume of organic matter. In general, the increasing pollution load promoted the accumulation of authigenic organic matter and exacerbated the pollution status of lakes (Zan et al., 2012). Indeed, Chaohu Lake now became one of the fi ve most polluted lakes in China. To alleviate the eutrophication of the lake, the local government launched the “Water Diversion Project” in 2012, in which Changjiang (Yangtze) River water is introduced into Chaohu Lake through Zhaohe River to shorten water cycle of the lake and alleviate eutrophication.
At present, most of the research projects on Chaohu Lake focus on heavy metals (Kong et al., 2015), nutrient changes (Liu et al., 2012), eutrophication (Huang et al., 2013), and control strategies (Shang and Shang, 2005). The bacterial community structure of the lake remains poorly understood. A previous study has shown that the bacterial community status is closely related to water quality and that changes in bacterial structure respond well to water pollution load (Zhang et al., 2015). Therefore, understanding the structural characteristics of bacterial communities is the key to study the pollution of Chaohu Lake. Although relationship between bacterial community structure and environmental factors in eutrophic shallow lakes has been widely investigated, studies on the efference ects of nutrients on the sediment fl ora structure under continuous seasonal changes are scarce. In this study, we chose Nanfei River and Zhaohe River estuaries as test sites, the water and sediment were sampled in spring, summer, autumn, and winter and analyzed. With rapid development in molecular biology, high-throughput sequencing technology has been widely used in environmental microbiology research (Huber et al., 2007). Compared with traditional monoclonal technology, DGGE (denaturing gradient gel electrophoresis), RFLP (restriction endonuclease fragment length polymorphism), and other approaches, highthroughput sequencing technology has obvious fl ux advantages and very high resolution, facilitating more comprehensive analysis with more details of the bacterial community structure of a sample.
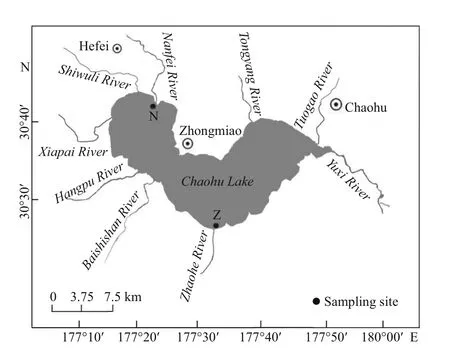
Fig.1 Map of the sampling sites in Chaohu Lake, China
In this study, 16S rRNA gene sequencing was performed, and the bacterial community structures were analyzed for the sediments of Nanfei River estuary and Zhaohe River estuary. Relationship between metabolic function of bacteria and environmental factors was clarifi ed to reveal the complexity of the lake ecology. The driving force behind the composition of water and sediment bacterial communities was determined for improving water quality and the ecological environment of the lake.
2 MATERIAL AND METHOD
2.1 Sampling sites and sample collection
The Nanfei River is located in the northwest corner of Chaohu Lake, by which many cities and counties situated with dense populations. Thus, the river estuary is the most polluted area of Chaohu Lake. The middle estuary area was selected for the study site (Site N) (Fig.1). The other site is located in the middle part of the Zhaohe River estuary (Site Z). As an artifi cial river, Zhaohe River intakes water from Changjiang River as a link between Changjiang River and Chaohu Lake. Four sets (seasonal) of water (NW8, NW11, NW2, and NW5) and sediment (NS8, NS11, NS2, and NS5) were sampled from Nanfei River estuary and four sets of water (ZW8, ZW11, ZW2, and ZW5) and sediment (ZS8, ZS11, ZS2, and ZS5) samples from Zhaohe River estuary. One water sample and one sediment sample were collected at a time in each site. The corresponding sampling times were August (summer) and November (autumn) 2016, and February (winter) and May (spring) 2017. For each site, surface sediment samples (<5 cm deep) were collected using a 1/16 m2Petersen grab sampler. Sediments of three separate grabs were homogenized as a composite sample, and three replicate sediments from each site were obtained. An aliquot of each composite sample was stored in a 15-mL sterile centrifuge tube at -80°C until freeze-drying and subsequent DNA extraction. Water was sampled approximately 50 cm below the surface using a vertical sampler and repeated three times; the samples were collected in plastic bottles pre-washed with deionized water.
2.2 Physicochemical analysis
At each site, pH, water temperature (Temp) and dissolved oxygen (DO) were measured on-site using a multi-parameter water quality analyzer (YSI 6600V2, USA). Chemical analyses, comprising total nitrogen (TN), total phosphorus (TP), and total organic carbon (TOC), were performed using standardized methods (Ruban et al., 2001) in triplication with a standard deviation lower than 5%.
2.3 DNA extraction and PCR amplifi cation
Bacterial genomic DNA of the 16 samples was extracted using the Fast DNA Spin Kit for Soil kit (MP Biomedicals, Fountain Pkwy. Solon, OH, USA). The concentration and purity of the genomic DNA were measured with an ultraviolet spectrophotometer (RS232G), and the integrity of the DNA sample was assessed by 0.8% agarose gel electrophoresis at a voltage of 120 V and an electrophoresis time of approximately 20 min. The extracted DNA stock wa s diluted to 20 ng/μL and used as a PCR template for sequencing of the highly variable V4 region of the bacterial 16S rRNA gene of approximately 400 bp in length. A universal primer for the V4 region was used (515F GTGCCAGCMGCCGCGGTAA/907R CCGTCAATTCMTTTRAGTTT) (Lane, 1991; Caporaso et al., 2011). The program was as follows: re-denaturation at 94°C for 5 min, denaturation at 50°C; annealing at 94°C for 30 s, 50°C for 30 s, and 72°C for 1 min for 27 cycles; 72°C for 5 min; hold at 4°C. The product was purifi ed using AXYGEN’s gel recovery kit. The PCR amplifi cation product was subjected to fl uorescence quantifi cation according to the preliminary quantitative results of electrophoresis with a Quant-iT PicoGreen dsDNA Assay Kit (Life Technologies cat, P11496) and a microplate reader (BioTek, FLx800). Based on the results, each sample was then mixed according to the amount of sequencing required and sent to Shanghai Personal Biotechnology Co., Ltd. for Illumina MiSeq high-throughput sequencing.
2.4 Statistical analysis
The alpha-diversity indices (Chao 1 index, Shannon index, Simpson index) of the 16 samples were calculated by software Mothur. Composition analysis was conducted at phylum and class levels. In order to assess the difference erence of bacterial community composition among samples of difference erent locations (Nanfei River estuary and Zhaohe River estuary) and sample types (water and sediment), analysis of similarities (ANOSIM) was performed based on Bray-Curtis dissimilarity with 9999 permutations using R package “vegan”. The NMDS was conducted in R with the vegan package. Bacteria of the dominant phylum and class in each sample were evaluated. To examine relationships between bacterial community structure and environmental parameters, redundancy analysis (RDA) was performed using CANOCO 4.5 software (Ter Braak and Smilauer, 2002).
3 RESULT
3.1 Physicochemical characteristics of samples
Major geographical and physiochemical characteristics of the lake sediments are summarized in Supplementary Table S1. Across the sampling sites, all water samples presented a slightly to moderately alkaline pH (7.68-8.49, average 8.14), and the temperature of the water in summer and winter was approximately 32°C and 3°C, respectively. The DO content in the water of the Nanfei River estuary and Zhaohe River estuary was highest in winter, at 11.95 mg/L and 11.40 mg/L, respectively. The TN and TP contents in each group showed seasonal changes. The range of TN in the water and sediment samples was 0.79-2.02 mg/L and 1.29- 2.15 g/kg, respectively, and that of TP was 0.06-0.27 mg/L and 0.17-1.56 g/kg, respectively. The TOC content in the sediments ranged 8.62-12.41 g/kg for the Nanfei River estuary and 1.49-3.94 g/kg for the Zhaohe River estuary, respectively; the TOC content in the two estuaries was signifi cantly difference erent ( P <0.05).
3.2 Biodiversity analysis
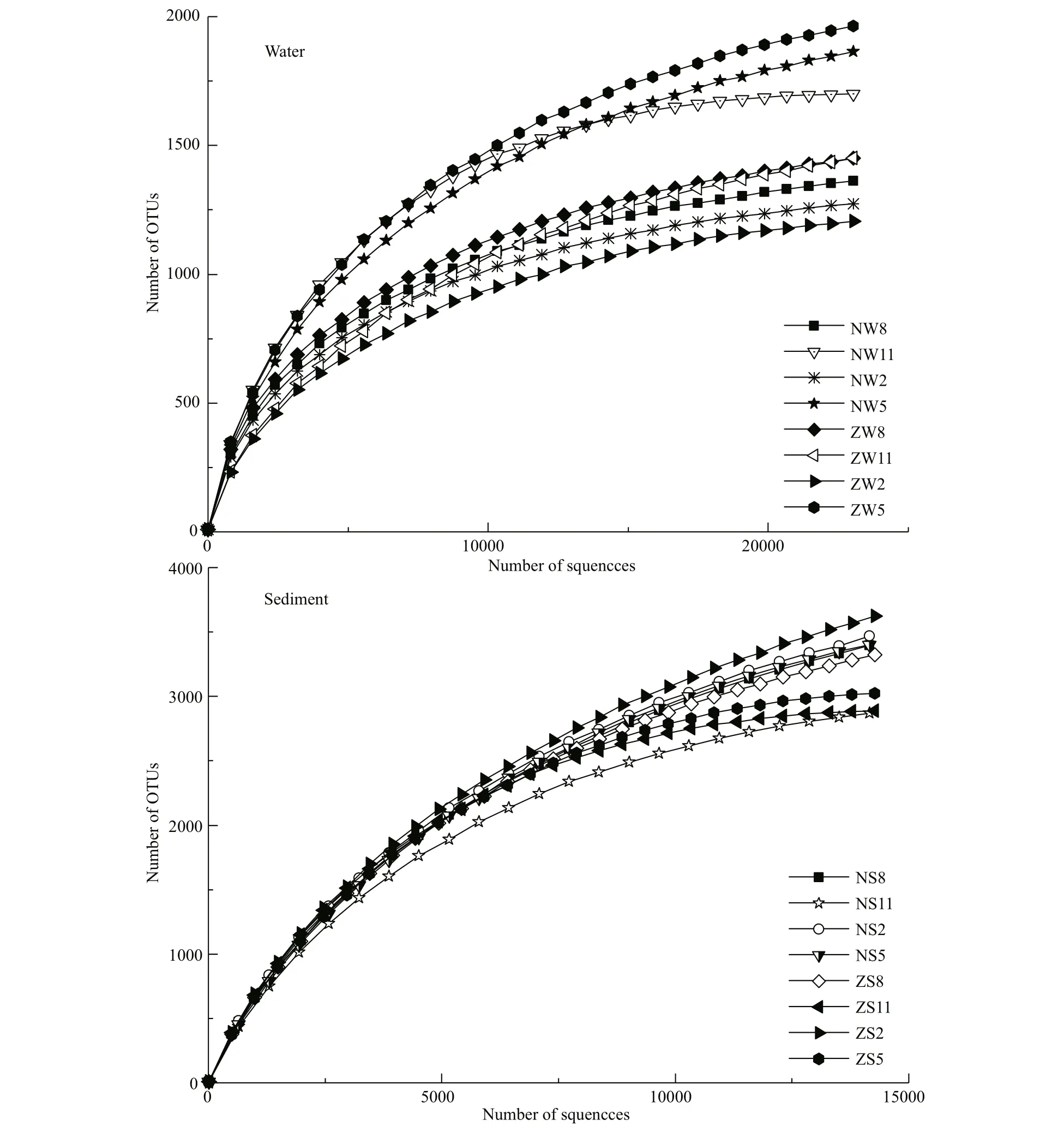
Fig.2 Rarefaction curves of OTUs clustered at 97% sequence identity across 16 samples from the Nanfei River and Zhaohe River estuaries
A total of 298 512 high-quality sequences were available from the 16 samples for subsequent analysis, with random sampling of the sequence for each. Rarefaction curves were constructed using the number of sequences and the number of OTUs between the difference erent samples (Fig.2). At 97% similarity level, the rarefaction curves of most samples reached saturation. A few samples were not completely saturated but approached the saturation state, indicating that most of the information from the samples was obtained and refl ected the structural composition of the bacterial community in the sediment. Diversity indexes (Chao 1 and Shannon) for the sediment samples were signifi cantly higher than those for the water samples ( P <0.05) (Table 1). Among them, ZS2 had the most diverse bacterial population (Shannon=10.61; Chao 1=5 135.74), and there was no signifi cant difference erence between the two sampling points. The seasonal variation trend for the Chao 1 index was the same among the water samples, and the sediment sample Chao 1 index results exhibited similar seasonal trends. In addition, ANOSIM indicated that abundant bacterial community composition significantly ( P <0.05) difference ered among the sediments samples in Zhaohe estuary (Supplementary Table S2).
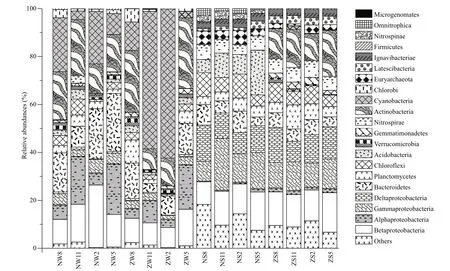
Fig.3 Relative abundances of the top 10 phyla or sub-phyla in each sample (a total of 21 phyla or classes for all 16 samples) from the Nanfei River and Zhaohe River estuaries
3.3 Analysis of dominant bacterium in the water and sediment of Chaohu Lake
Figure 3 shows the distribution of dominant bacteria in the samples at phylum level. Proteobacteria (34.02±9.86)%, Actinobacteria (12.86±9.48)%, Cyanobacteria (12.07±20.10)%, Bacteroidetes (9.85±6.29)%, Chlorofl exi (4.94±2.98)%, Planctomycetes (4.52±2.57)%, Acidobacteria (3.54±5.07)%, Nitrospirae (3.15±2.99)%, Verrucomicrobia (1.31±1.17)%, and Gemmatimonadetes (1.17±0.98)% were common dominant bacteria in water and sediment samples. Chlorobi (1.81±1.78)%, Firmicutes (0.15±0.20)%, and Microgenomates (0.05±0.12)% were dominant bacteria in water samples only, whereas Euryarchaeota (3.33±1.73)%, Ignavibacteriae (3.20±0.33)%, Latescibacteria (2.51±0.99)%, Nitrospinae (1.25±1.45)%, and Omnitrophica (0.79±1.08)% were dominant in sediment samples only.
The bacterial community composition of the water and sediment samples showed similarity at phylum level but signifi cant difference erences at genus level (Figs.4 & 5). For example, in the water samples, unclassifi ed Family I (23.32±22.59)% was the most abundant bacterium, followed by hgcI clade (13.88±5.84)%,
unclassifi ed LD12 freshwater group (6.12±6.74)%, CL500-29 marine group (3.51±2.54)%, unclassifi ed Comamonadaceae (2.94±3.72)%, unclassifi ed Chloroplast (2.90±6.34)% and Flavobacterium (2.56±4.34)%. Among sediment samples, unclassifi ed Xanthomonadales incertae sedis (7.35±4.12)% was the most abundant bacterium, followed by unclassifi ed Anaerolineaceae (4.61±1.38)%, unclassifi ed Nitrospiraceae (3.92±0.96)%, unclassifi ed 43F-1404R (3.57±2.19)%, unclassifi ed Subgroup_ 22 (3.34±0.91)%, Subgroup 6 (2.97±2.02)%, and unclassifi ed SC-I-84 (2.85±1.73)%. Overall, the bacterial community of sediment samples displayed higher diversity and uniformity compared to the water samples.
3.4 Factors infl uencing the bacterial communities

Fig.4 Relative abundances of the top 20 genera in water samples (a total of 48 genera or unclassifi ed genus) from the Nanfei River and Zhaohe River estuaries
Bacterial abundance, community succession, and functional activity respond quickly to changes in nutrient salts, hydrology, and other environmental factors in lakes (Paerl et al., 2003; Shade et al., 2011). RDA analysis was used to determine the efference ects of environmental factors on bacterial community structure, and the physicochemical parameters that signifi cantly afference ect the bacterial community were selected using the forward selection method in CANOCO 4.5 (Ter Braak and Smilauer, 2002). Figure 6 illustrates the relationship between bacterium and environmental factors in the water. Betaproteobacteria, Deltaproteobacteria, Gammaproteobacteria, Bacteroidetes, Chlorofl exi, Acidobacteria, and Actinobacteria were correlated positively with TN and T and negatively with TP and DO; Cyanobacteria and Microgenomates showed the opposite trends. Alphaproteobacteria correlated positively with TP and TN and negatively with DO. Relationship between bacterium and environmental factors in sediments is presented in Fig.7. As shown, the relative abundances of Betaproteobacteria, Alphaproteobacteria, Deltaproteobacteria, Actinobacteria, Cyanobacteria, Latescibacteria, and Verrucomicrobia were correlated negatively with TP, TN, and TOC, but Acidobacteria, Gammaproteobacteria, Euryarchaeota and Omnitrophica were correlated positively with TP, TN, and TOC. These results suggest that environmental factors have difference erent positive or negative implications for the distributions of these bacteria.
4 DISCUSSION
Nanfei River and Zhaohe River estuaries are representative areas for studying the bacterial community structure of the water and sediment in Chaohu Lake. Our data show temporal and spatial difference erences between physical and chemical indicators for the water and sediment samples (Supplementary Table S1), which results from the combination of hydrodynamic disturbance, molecular difference usion, and bioturbation. Phosphorus (Sun et al., 2010) and other contaminants (Yu et al., 2014) are converted to inorganic substances by mineralization and degradation via the activities of microorganisms and are released into the lake by pore water under certain conditions. Overall, the nitrogen and phosphorus contents in the water at the sampling sites increased. The TP, TN, and TOC contents in the sediments of Nanfei River estuary were signifi cantly higher than those of Zhaohe River estuary. Among them, the seasonal variation of TP content in Nanfei River estuary was 0.97-1.56 g/kg, and the seasonal variation in Zhaohe River estuary was 0.17-0.41 g/kg. The TP content displayed a signifi cant decreasing trend from west to east in Chaohu Lake sediment (Ding et al., 2014) along with the development of agricultural and phosphorus deposits around the lake area. Continuous mining may be the main reason for the high TP in the western region of the lake (Zan et al., 2011).

Fig.5 Relative abundances of the top 20 genera in sedimentary samples (a total of 38 genera or unclassifi ed genus) from the Nanfei River and Zhaohe River estuaries
Species diversity is a prerequisite for maintaining the normal function of ecosystems. The richness and diversity of bacterial communities play an important role in nutrient salt cycling, organic matter degradation, heavy metal transformation, and greenhouse gas emissions in lake ecosystems (Ansola et al., 2014). NMDS (Supplementary Fig.S1) show that most of the water samples and sediment samples have similar bacterial community composition in the same season at the same sampling point. For example, NS5 and NW5, ZS8 and ZW8, ZS5 and ZW5 have similar bacterial community composition. Similar temperature conditions may be the main reason for the similar composition of bacterial communities in water and sediments, and the interaction of water and sediments creates similar nutrient conditions for bacterial communities. Previous studies have also shown that nutrients (Xing and Kong, 2007) and temperature (Deng et al., 2007) have an important infl uence on the composition of bacterial communities. In this study, the Chao 1 index and Shannon index values of the sediment samples were higher than those of the water samples, likely because there are much higher levels of nutrients in sediments than in water, providing a suitable breeding environment for the growth and reproduction of bacteria and promoting the growth of a variety of bacteria, which in turn leads to a more complex bacterial community structure (Bai et al., 2012). In addition, because the sampling points were located in estuaries, there is a certain fl uidity to the water fl ow. Removing a large number of planktonic bacteria during the process of water body renewal may reduce the diversity of the bacterial community of the water sample.
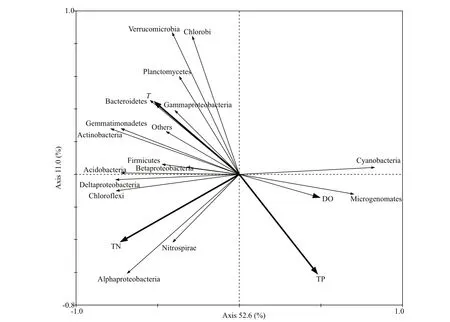
Fig.6 Water analysis (RDA) of bacterial communities and correlation between relative abundances of major bacterium and main water parameters
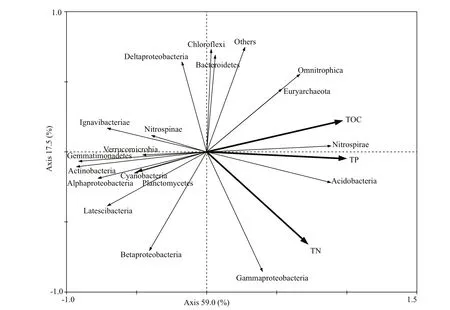
Fig.7 Sediment analysis (RDA) of bacterial communities and correlation between relative abundances of major bacterium and main sediment parameters
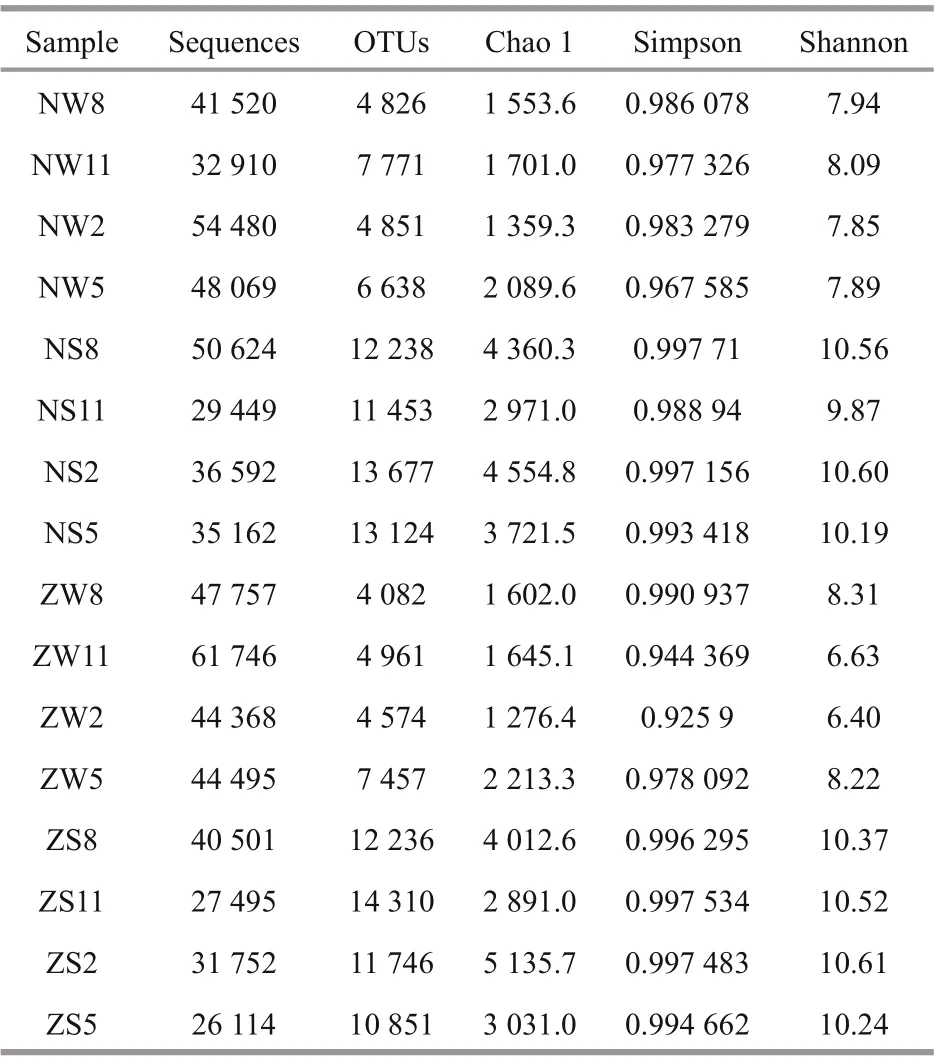
Table 1 Diversity of bacterial 16S rRNA gene libraries of 16 samples in Chaohu Lake
Changes in bacterial community composition are one of the most sensitive indicators refl ecting lake eutrophication; thus, understanding the composition of bacterial communities facilitates a better understanding of the metabolic processes of aquatic ecosystems (Dillon et al., 2009). In this study, the temporal distribution of most bacterial communities in sediment samples varied widely ( P <0.05). Among 16 samples, Proteobacteria was the most advantageous bacteria in all samples. Most Proteobacteria have a strong ability to decompose difference erent materials and can be widely used to metabolize various types of organic matter (Xue et al., 2018), thus playing a vital role in the nutrient cycle of lakes. As depicted in Fig.3, seasonal variation in the relative abundance of Betaproteobacteria (13.61±4.21)% was minimal, and this phylum was dominant in both water and sediment samples. However, the advantage of this bacterium in the sediment samples was more evident than in the water samples, indicating that Betaproteobacteria can adapt well in difference erent environments and resist temperature fl uctuations (Newton et al., 2011). Betaproteobacteria serve as decomposers of complex organisms to produce nutrients such as ammonia and methane (Dang et al., 2010), which is consistent with the positive correlation between Betaproteobacteria and TN in RDA. As the Nanfei River estuary has a high TN content, a suき cient amount of nutrients may be the main reason for the dominance of this phylum in the estuary (Kielak et al., 2016). In this study, a large number of Alcaligenaceae (2.45±1.15)% belonging to Betaproteobacteria were found in sediments, species that are widely distributed in various habitats, from animals and humans to soil, sewage and sludge (Dam et al., 2009). The metabolic patterns of Alcaligenaceae are also diverse (Ghosh et al., 2005), and studies have shown that this family carries a large complement of plasmids (Riccio et al., 2001) that contain genes involved in biodegradation conferring Alcaligenaceae with functional features related to biodegradation and biogeochemistry (Wyndham et al., 1988).
Gammaproteobacteria (8.12±5.59)% and Deltaproteobacteria (6.24±5.97)% were also dominant among Proteobacteria, with higher relative abundance in sediment samples than in water samples. Gammaproteobacteria can participate in the decomposition of various organic carbons and organic phosphorus (Čanković et al., 2017). Unclassifi ed Xanthomonadales incertae sedis, a member of Gammaproteobacteria with the ability to degrade aromatics (Xiao and Ni, 2013), also showed high relative abundance in this study. Overall, analysis of this taxon will help to mitigate the pollution of Chaohu Lake and maintain ecological balance. The study of the bacteria contributes to the removal of pollution and the maintenance of the ecological balance in Chaohu Lake. Deltaproteobacteria includes basic aerobic slime bacteria and some obligate anaerobic species, which degrade organic matter in the sediment via the oxidation and reduction of sulfur compounds (Čanković et al., 2017). In this study, unclassifi ed Anaeromyxobacter (0.37±0.41)% was one of the important members of Deltaproteobacteria, which was only found in Nanfei River estuary. Subclass Anaeromyxobacter can utilize a variety of electron acceptors, including heavy metals complexed as nitrates, ferric iron, and manganese oxide (Thomas et al., 2009). Due to the dependence of Anaeromyxobacter on heavy metals and nitrates, observing the distribution of these species can help to elucidate the distribution and migration of pollutants in Chaohu Lake. In addition, Geothermobacter, an iron-reducing bacterium, was enriched (0.71±0.59)% in the sediment of Nanfei River estuary. Under anaerobic conditions, Geothermobacter can utilize acetic acid, formic acid, and succinic acid. As an electron donor, Fe3+acts as a terminal electron acceptor, reducing various toxic heavy metals, degrading and consuming organic and inorganic contaminants and eventually reducing Fe3+to Fe2+(Lovley et al., 2011). The relative abundance of Geothermobacter in Nanfei River estuary was much higher than that in Zhaohe River estuary, indicating that sediments in Nanfei River estuary may contain higher concentrations of heavy metals and organic acids.
Alphaproteobacteria is a major group in highsalinity environments, but the relative abundance of Alphaproteobacteria (10.96±7.61)% in our water samples was high. RDA showed that Alphaproteobacteria correlated positively with TN and TP in water, which is consistent with the degradation of complex aromatic compounds and the anaerobic oxidation of ammonium and nitrite (Kaluzhnaya et al., 2011). The anthropogenic emissions of organic pollutants around the river are the main factors that promote the enrichment of Alphaproteobacteria in the Nanfei River estuary. In this study, the LD12 freshwater clade was the most abundant bacterium of Alphaproteobacteria. It has been reported that LD12 is the main genus in marine and freshwater systems (Henson et al., 2018), and the presence of this taxon in a freshwater system (Logares et al., 2010; Eiler et al., 2014) indicates changes in its ecological development. It has successfully adapted and migrated from the saltwater environment to the freshwater environment.
Bacteroidetes generally have the ability to settle and decompose hydrocarbons (Kasai et al., 2001) and are widely found in eutrophic lake waters (Tang et al., 2015) and sediments (Newton and McMahon, 2011). In our study, RDA showed that Bacteroidetes correlated positively with TOC, which is consistent with the functional characteristics of these bacteria. Studies have also shown that a large number of Bacteroidetes are often present in oil-contaminated environments, which may be due to their ability to secrete polysaccharide mucus, facilitating movement on smooth surfaces (Gao et al., 2014). Compared with the water samples in Zhaohe River in the lake area, the relative abundance of Bacteroidetes in Nanfei River estuary was higher (Fig.3), indicating that there may be a more severe condition of hydrocarbon pollution in Nanfei River estuary. Many branches of the Bacteroidetes family were identifi ed, among which the relative abundance of Flavobacterium at NW2 (6.17%) and NW5 (12.01%) was signifi cantly higher than at other sites. Studies have shown that Flavobacterium is a common aerobic denitrifying bacterium with heterotrophic nitrifi cation and the ability to metabolize refractory organics under conditions of suき cient dissolved oxygen (Zhou et al., 2007). Proper temperature, adequate dissolved oxygen and abundant nutrients may be the main reason for the predominance of Flavobacterium at NW2 and NW5. In addition, a large number of Bacteroidetes members with a relative abundance of less than 2% were found in the sediment, such as unclassifi ed Bacteroidetes_vadinHA17 (0.35±0.50)%, unclassifi ed Lentimicrobiaceae (1.82±0.66)%, unclassifi ed Cytophagaceae (1.45±1.28)% and unclassifi ed Saprospiraceae (0.38±0.59)%.
Currently, few Chlorofl exi lineages been cultured and identifi ed, and the specifi c functions of Chlorofl exi within the context of ecosystems remain unknown. Nonetheless, studies have shown that the bacterium of this genus are widely distributed (Edmonds-Wilson et al., 2015), with a variety of metabolic types, including anaerobic phototrophs, anoxygenic phototrophs, obligate anaerobic heterotrophs, and anaerobic halorespirers. Chlorofl exi can even exhibit a prey-specifi c growth response (Yamada and Sekiguchi, 2009). Hug et al. (2013) noted that the presence of Chlorofl exi is often associated with uranium-contaminated aquifers. Therefore, the detection of a higher abundance of Chlorofl exi in water samples indicates that there may be some uranium pollution in Nanfei River and Zhaohe River estuaries. RDA indicated that the abundance of Chlorofl exi correlated negatively with TP (Fig.6), which is consistent with the fi ndings of Song et al. (2012). In our study, unclassifi ed Anaerolineaceae was the most dominant member of Chlorofl exi in sediments, with a relative abundance ranging 3.27%-7.02%. Other studies on relative abundance have shown that Anaerolineaceae functions in the degradation of long-chain n-alkane species (N5-Z0) and cellular amino acid materials (Lv et al., 2014; Liang et al., 2016), accelerating the degradation of organic carbon compounds and cells in sediments and helping to restore and purify the ecosystem of Chaohu Lake.
The phylum Actinobacteria comprises a wide variety of members found in freshwater environments (Parveen et al., 2011). Haukka et al. (2006) reported that Actinobacteria prefer to live in a noneutrophic environment. Accordingly, Actinomycetes were not detected in the sediments of the Nanfei River estuary, but the higher relative abundance in other samples confi rms that the eutrophication of the Nanfei River estuary is serious. Our data showed hgeI clade and CL500-29 marine group to be representative of Actinobacteria. CL500-29 marine group has a strong capacity for absorbing carbohydrates and nitrogenrich compounds and well tolerates low-DO environments (Lindh et al., 2015); hgeI clade has similar characteristics (Ghylin et al., 2014). Among them, the relative abundance of CL500-29 marine group in Nanfei River estuary and Zhaohe River estuary was highest in summer, which may be caused by the synergy of low DO, high T and TN and some unknown driving factors.
Cyanobacteria (23.86±23.40)% species are mainly distributed in water samples; they are more common in eutrophic alkaline waters with higher nitrogen and phosphorus contents and can be used as an indicator of the eutrophication of water bodies (Chen et al., 2013). Monitoring Cyanobacteria can provide a theoretical basis for the control of algal blooms in Chaohu Lake and create conditions for the restoration of the water ecosystem of this lake (Zhao et al., 2002). In RDA, TP and DO showed a positive correlation with Cyanobacteria; thus, increases in TP and DO concentrations in a certain range have a positive regulatory efference ect on the relative abundance of Cyanobacteria. Cyanobacterial blooms usually occur in summer and rarely occur under 15°C (Whitton and Potts, 2007). Surprisingly, the relative abundance of Cyanobacteria in Zhaohe River estuary was over 57% in both autumn and winter, which indicates that in addition to temperature factors, other environmental factors in Chaohu Lake play a major role in regulating the distribution of this phylum. Among Cyanobacteria phylum, unclassifi ed Family I and Microcystis (2.55±3.07)% were the most dominant at the two test points. The latter comprises a group of non-nitrogenfi xing bacteria that degrade organic compounds such as nitrogen and phosphorus (Nalewajko and Murphy, 2001). In this study, the relative abundance of Microcystis varied seasonally, indicating that temperature is an important factor afference ecting the distribution of these bacteria (Robarts and Zohary, 1987). Spatially, the relative abundance of Microcystis in Nanfei River estuary was higher than that in Zhaohe River estuary, which may be due to the higher contents of nitrogen and phosphorus compounds in the former, which provides suき cient nutrition for the growth and reproduction of Microcystis.
Acidobacteria, Euryarchaeota, and Ignavibacteriae also decompose organic carbon (Mehta and Baross, 2006; Podosokorskaya et al., 2013), and Omnitrophica (tentatively classifi ed as OP3) and Latescibacteria (tentatively classifi ed as WS3) are magnetotactic bacteria capable of synthesizing magnetite (Fe3O4) from cells or pyrite (Fe3S4) crystal particles (Lin and Pan, 2015). In addition, Nitrospirae and Planctomycetes (Schneider et al., 2013) can to some extent decompose nitrogen-containing compounds in sediments. The fact that the organic matter content in sediments is much higher than that in water shall be the main reason why these bacteria are concentrated in sediments.
5 CONCLUSION
Chaohu Lake is a large shallow, eutrophic freshwater lake in China. Due to historical reasons and human activities, Chaohu Lake water pollution has become increasingly serious. In this study, 16S rRNA high-throughput sequencing technology was used to investigate the bacterial community structure of water and sediments in Nanfei River estuary and Zhaohe River estuary, providing a theoretical basis for the treatment of eutrophic water in Chaohu Lake. In this study, the physical and chemical indicators of TN, TOC, TP, and other parameters were signifi cantly higher in Nanfei River estuary than in Zhaohe River estuary. Moreover, the discharge of sewage around Nanfei River and the mining of phosphate rock have increased the pollution of the lake area. The water of Changjiang River was introduced into Chaohu Lake via Zhaohe River. Compared with Nanfei River estuary, the water source of Zhaohe River estuary is cleaner. Our data show that Betaproteobacteria is a unique bacterial community with stable distribution in every sample. Overall, the observed changes in the water and sediment bacterial community have spatial specifi city and seasonal characteristics. The bacterial community structure of Nanfei River estuary and Zhaohe River estuary difference ered greatly at the genus level, and the nutritional status may be a key force driving the lake and sediment bacterial communities. In addition, a large number of unidentifi able genera were found in the study, indicating that the bacterial community in Chaohu Lake sediments is very diverse.
6 DATA AVAILABILITY STATEMENT
All sequence data were deposited in the National Center for Biotechnology Information (NCBI) Sequence Read Archive with accession No. SRP217654.
7 ACKNOWLEDGMENT
Anonymous reviewers are acknowledged for their constructive comments and helpful suggestions.
8 CONFLICTS OF INTEREST
The authors declare that they have no confl icts of interest.
References
Ansola G, Arroyo P, De Miera L E S. 2014. Characterisation of the soil bacterial community structure and composition of natural and constructed wetlands. Sci. Total. Environ., 473- 474: 63-71.
Bai Y H, Shi Q, Wen D H, Li Z X, Jefference erson W A, Feng C P, Tang X Y. 2012. Bacterial communities in the sediments of Dianchi Lake, a partitioned eutrophic waterbody in China. PLoS One, 7(5): e37796.
Čanković M, Petrić I, Marguš M, Ciglenečki I. 2017. Spatiotemporal dynamics of sulfate-reducing bacteria in extreme environment of Rogoznica Lake revealed by 16S rRNA analysis. J. Marine Syst., 172: 14-23.
Cao Y M, Zhang E L, Cheng G J. 2014. A primary study on relationships between subfossil chironomids and the distribution of aquatic macrophytes in three lowland fl oodplain lakes, China. Aquat. Ecol., 48(4): 481-492.
Caporaso J G, Lauber C L, Walters W A, Berg-Lyons D, Lozupone C A, Turnbaugh P J, Fierer N, Knight R. 2011. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. P roc. Natl. Acad. Sci. USA, 108(S1): 4 516-4 522.
Chen Z B, Zhou Z Y, Peng X, Xiang H, Xiang S N, Jiang Z X. 2013. Efference ects of wet and dry seasons on the aquatic bacterial community structure of the Three Gorges Reservoir. World J. Microb iol. Biot echnol., 29(5): 841-853.
Dam B, Ghosh W, Gupta S K D. 2009. Das Gupta. Conjugative Type 4 secretion system of a novel large plasmid from the chemoautotroph Tetrathiobacter kashmirensis and construction of shuttle vectors for Alcaligenaceae. Appl. Environ. Microb iol., 75(13): 4 362-4 373.
Dang H Y, Li J, Chen R P, Wang L, Guo L Z, Zhang Z N, Klotz M G. 2010. Diversity, abundance, and spatial distribution of sediment ammonia-oxidizing Betaproteobacteria in response to environmental gradients and coastal eutrophication in Jiaozhou Bay, China. Appl. Environ. Microb iol., 76(14): 4 691-4 702.
Deng D G, Xie P, Zhou Q, Yang H, Cuo L G. 2007. Studies on temporal and spatial variations of phytoplankton in Lake Chaohu. J. Integr. Plant Biol., 49(4): 409-418.
Dillon J G, McMath L M, Trout A L. 2009. Seasonal changes in bacterial diversity in the Salton Sea. Hydrobiologia, 632(1): 49-64.
Ding W, Zhu R B, Hou L J, Wang Q. 2014. Matrix-bound phosphine, phosphorus fractions and phosphatase activity through sediment profi les in Lake Chaohu, China. Environ. Sci. Proc ess Impacts, 16(5): 1 135-1 144.
Edmonds-Wilson S L, Nurinova N I, Zapka C A, Fierer N, Wilson M. 2015. Review of human hand microbiome research. J. Dermatol. Sci., 80(1): 3-12.
Eggleton J, Thomas K V. 2004. A review of factors afference ecting the release and bioavailability of contaminants during sediment disturbance events. Environ. Int., 30(7): 973-980.
Eiler A, Zaremba-Niedzwiedzka K, Martínez-García M, McMahon K D, Stepanauskas R, Andersson S G, Bertilsson S. 2014. Productivity and salinity structuring of the microplankton revealed by comparative freshwater metagenomics. Environ. Microb iol., 16(9): 2 682-2 698. Fernández-Luqueño F, Valenzuela-Encinas C, Marsch R, Martínez-Suárez C, Vázquez-Núñez E, Dendooven L. 2011. Microbial communities to mitigate contamination of PAHs in soil—possibilities and challenges: a review. Environ. Sci. Pollut. R es., 18(1): 12-30.
Fuhrman J A, McCallum K, Davis A A. 1993. Phylogenetic diversity of subsurface marine microbial communities from the Atlantic and Pacifi c Oceans. Appl. Environ. Microb iol., 59(5): 1 294-1 302.
Gao X Y, Ming H X, Chen J Y, Li J Y, Han J L, Lin F A, Fan J F. 2014. Bacterial community in Dalian Bay petroleum pollution sediments. Acta Oceanol. Sin., 36(6): 58-66. (in Chinese with English abstract)
Ghosh W, Bagchi A, Mandal S, Dam B, Roy P. 2005. Tetrathiobacter kashmirensis gen. nov., sp. nov., a novel mesophilic, neutrophilic, tetrathionate-oxidizing, facultatively chemolithotrophic Betaproteobacterium isolated from soil from a temperate orchard in Jammu and Kashmir, India. Int. J. Syst. Evol. Micr obiol., 55(5): 1 779-1 787.
Ghylin T W, Garcia S L, Moya F, Oyserman B O, Schwientek P, Forest K T, Mutschler J, Dwulit-Smith J, Chan L K, Martinez-Garcia M, Sczyrba A, Stepanauskas R, Grossart H P, Woyke T, Warnecke F, Malmstrom R, Bertilsson S, McMahon K D. 2014. Comparative single-cell genomics reveals potential ecological niches for the freshwater acI Actinobacteria lineage. ISME J., 8(12): 2 503-2 516.
Haukka K, Kolmonen E, Hyder R, Hietala J, Vakkilainen K, Kairesalo T, Haario H, Sivonen K. 2006. Efference ect of nutrient loading on bacterioplankton community composition in Lake Mesocosms. Microb. Ecol., 51(2): 137-146.
Henson M W, Lanclos V C, Faircloth B C, Thrash J C. 2018. Cultivation and genomics of the fi rst freshwater SAR11 (LD12) isolate. ISME J., 12(7): 1 846-1 860.
Huang J, Zhan J Y, Yan H M, Wu F, Deng X Z. 2013. Evaluation of the impacts of land use on water quality: a case study in The Chaohu Lake Basin. Scientifi c World J., 2013: 329187.
Huber J A, Mark Welch D B, Morrison H G, Huse S M, Neal P R, Butterfi eld D A, Sogin M L. 2007. Microbial population structures in the deep marine biosphere. Sci ence, 318(5847): 97-100.
Hug L A, Castelle C J, Wrighton K C, Thomas B C, Sharon I, Frischkorn K R, Williams K H, Tringe S G, Banfi eld J F. 2013. Community genomic analyses constrain the distribution of metabolic traits across the Chlorofl exi phylum and indicate roles in sediment carbon cycling. Microbiome, 1(1): 22.
Kaluzhnaya O V, Itskovich V B, McCormack G P. 2011. Phylogenetic diversity of bacteria associated with the endemic freshwater sponge Lubomirskia baicalensis. World J. Microb iol. Biot echnol., 27(8): 1 955-1 959.
Kasai Y, Kishira H, Syutsubo K, Harayama S. 2001. Molecular detection of marine bacterial populations on beaches contaminated by the Nakhodka tanker oil-spill accident. Environ. Microb iol., 3(4): 246-255.
Kielak A M, Barreto C C, Kowalchuk G A, van Veen J A, Kuramae E E. 2016. The ecology of Acidobacteria: moving beyond genes and genomes. Front. Microbiol., 7: 744.
Kong M, Dong Z L, Chao J Y, Zhang Y M, Yin H B. 2015. Bioavailability and ecological risk assessment of heavy metals in surface sediments of Lake Chaohu. China Environ. Sci., 35(4): 1 223-1 229. (in Chinese with English abstract)
Lane D J. 1991. 16S/23S rRNA sequencing. In: Stackebrandt E, Goodfellow M eds. Nucleic Acid Techniques in Bacterial Systematics. Wiley, New York. p.115-175.
Liang B, Wang L Y, Zhou Z C, Mbadinga S M, Zhou L, Liu J F, Yang S Z, Gu J D, Mu B Z. 2016. High frequency of Thermodesulfovibrio spp. and Anaerolineaceae in association with Methanoculleus spp. in a long-term incubation of n-alkanes-degrading methanogenic enrichment culture. Front. Microbiol., 7: 1 431.
Lin W, Pan Y X. 2015. A putative greigite-type magnetosome gene cluster from the candidate phylum Latescibacteria. Env iron. Mircobiol. Rep., 7(2): 237-242.
Lindh M V, Lefébure R, Degerman R, Lundin D, Andersson A, Pinhassi J. 2015. Consequences of increased terrestrial dissolved organic matter and temperature on bacterioplankton community composition during a Baltic Sea mesocosm experiment. AMBIO, 44(S3): 402-412.
Liu E F, Shen J, Birch G F, Yang X D, Wu Y H, Xue B. 2012. Human-induced change in sedimentary trace metals and phosphorus in Chaohu Lake, China, over the past halfmillennium. J. Paleolimnol., 47(4): 677-691.
Liu L M, Yang J, Zhang Y Y. 2011. Genetic diversity patterns of microbial communities in a subtropical riverine ecosystem (Jiulong River, Southeast China). Hydrobiologia, 678(1): 113-125.
Logares R, Bråte J, Heinrich F, Shalchian-Tabrizi K, Bertilsson S. 2010. Infrequent transitions between saline and fresh waters in one of the Most Abundant Microbial Lineages (SAR11). Mol. Biol. Evol., 27(2): 347-357.
Lovley D R, Ueki T, Zhang T, Malvankar N S, Shrestha P M, Flanagan K A, Aklujkar M, Butler J E, Giloteaux L, Rotaru A E, Holmes D E, Franks A E, Orellana R, Risso C, Nevin K P. 2011. Geobacter: the microbe electric's physiology, ecology, and practical applications. Adv. Microb. Physiol., 59: 1-100.
Lv X F, Yu J B, Fu Y Q, Ma B, Qiu F Z, Ning K, Wu H F. 2014. A meta-analysis of the bacterial and archaeal diversity observed in wetland soils. Sci World J., 2014: 437684.
Mehta M P, Baross J A. 2006. Nitrogen fi xation at 92°C by a hydrothermal vent archaeon. Science, 314(5806): 1 783-1 786.
Meng P, Ma T. 2015. Discussion on the current situation, causes and ecological treatment of water pollution in Chaohu Lake. Res. Economi. Environ. Pro t., (1): 171-173. (in Chinese)
Mills H J, Martinez R J, Story S, Sobecky P A. 2004. Identifi cation of members of the metabolically active microbial populations associated with Beggiatoa species mat communities from Gulf of Mexico cold-seep sediments. Appl. Environ. Microb iol., 70(9): 5 447-5 458.
Nalewajko C, Murphy T P. 2001. Efference ects of temperature, and availability of nitrogen and phosphorus on the abundance of Anabaena and Microcystis in Lake Biwa, Japan: an experimental approach. Limnology, 2(1): 45-48.
Newton R J, Jones S E, Eiler A, McMahon K D, Bertilsson S. 2011. A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. R ev., 75(1): 14-49.
Newton R J, McMahon K D. 2011. Seasonal difference erences in bacterial community composition following nutrient additions in a eutrophic lake. Environ. Microb iol, 13(4): 887-899.
Paerl H W, Dyble J, Moisander P H, Noble R T, Piehler M F, Pinckney J L, Steppe T F, Twomey L, Valdes L M. 2003. Microbial indicators of aquatic ecosystem change: current applications to eutrophication studies. FEMS Microbiol. Ecol., 46(3): 233-246.
Parveen B, Reveilliez J P, Mary I, Ravet V, Bronner G, Mangot J F, Domaizon I, Debroas D. 2011. Diversity and dynamics of free-living and particle-associated Betaproteobacteria and Actinobacteria in relation to phytoplankton and zooplankton communities. FEMS Microbiol. Ecol., 77(3): 461-476.
Podosokorskaya O A, Kadnikov V V, Gavrilov S N, Mardanov A V, Merkel A Y, Karnachuk O V, Ravin N V, Bonch-Osmolovskaya E A, Kublanov I V. 2013. Characterization of Melioribacter roseus gen. nov., sp. nov., a novel facultatively anaerobic thermophilic cellulolytic bacterium from the class Ignavibacteria, and a proposal of a novel bacterial phylum Ignavibacteriae. Environ. Microbiol., 15(6): 1 759-1 771.
Riccio M L, Pallecchi L, Fontana R, Rossolini G M. 2001. In70 of plasmid pAX22, a blaVIM-1-containing integron carrying a new aminoglycoside phosphotransferase gene cassette. Antimicrob. Age nts Chemother., 45(4): 1 249-1 253.
Robarts R D, Zohary T. 1987. Temperature efference ects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. New Zeal. J. Mar. Fresh wat. Res., 21(3): 391-399.
Ruban V, López-Sánchez J F, Pardo P, Rauret G, Muntau H, Quevauviller P. 2001. Development of a harmonised phosphorus extraction procedure and certifi cation of a sediment reference material. J. Environ. Monit., 3(1): 121-125.
Schneider D, Arp G, Reimer A, Reitner J, Daniel R. 2013. Phylogenetic analysis of a microbialite-forming microbial mat from a hypersaline lake of the Kiritimati atoll, Central Pacifi c. PLoS One, 8(6): e66662.
Shade A, Read J S, Welkie D G, Kratz T K, Wu C H, McMahon K D. 2011. Resistance, resilience and recovery: aquatic bacterial dynamics after water column disturbance. Environ. Microbiol., 13(10): 2 752-2 767.
Shang G P, Shang J C. 2005. Causes and control countermeasures of eutrophication in Chaohu lake, China. Chin. Geogr. Sci., 15(4): 348-354.
Song H, Li Z, Du B, Wang G, Ding Y. 2012. Bacterial communities in sediments of the shallow Lake Dongping in China. J. Appl. Microbiol., 112(1): 79-89.
Sun Q Y, Ma X L, Yang G D, Chen Z, Wu H L, Xuan H X. 2010. Studies on nitrogen, phosphorus and organic matter in ponds around Chaohu Lake. Environ. Sci., 31(7): 1 510-1 515. (in Chinese with English abstract)
Tang X M, Li L L, Shao K Q, Wang B W, Cai X L, Zhang L, Chao J Y, Gao G. 2015. Pyrosequencing analysis of freeliving and attached bacterial communities in Meiliang Bay, Lake Taihu, a large eutrophic shallow lake in China. Can. J. Microbiol., 61(1): 22-31.
Ter Braak C J F, Smilauer P. 2002. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5). Wageningen University, Ithaca, NY, USA.
Thomas S H, Padilla-Crespo E, Jardine P M, Sanford R A, Löラ er F E. 2009. Diversity and distribution of Anaeromyxobacter strains in a uranium-contaminated subsurface environment with a nonuniform groundwater fl ow. Appl. Environ. Microb iol., 75(11): 3 679-3 687.
Whitton B A, Potts M. 2000. The Ecology of Cyanobacteria: Their Diversity in Time and Space. Kluwer Academic, Dordrecht, Boston.
Wyndham R C, Singh R K, Straus N A. 1988. Catabolic instability, plasmid gene deletion and recombination in Alcaligenes sp. BR60. Arch. Microbiol., 150(3): 237-243.
Xiao H H, Ni J R. 2013. Characterization microbial community structure in wastewater treatment plants (WWTPs) through 16S rRNA clone library. J. Bas ic Sci. Eng., 21(3): 522-531. (in Chinese with English abstract)
Xing P, Kong F X. 2007. Intra-habitat heterogeneity of environmental factors regulating bacterioplankton community composition in Lake Taihu, China. Aquat. Microb. Ecol., 48(2): 113-122.
Xue Y G, Liu F, Sun M. 2018. Community structure and infl uencing factors of bacterioplankton in spring in Zhushan Bay, Lake Taihu. Environ. Sci., 3 9(3): 1 151-1 158. (in Chinese with English abstract)
Yamada T, Sekiguchi Y. 2009. Cultivation of uncultured chlorofl exi subphyla: signifi cance and ecophysiology of formerly uncultured chlorofl exi ‘subphylum i’ with natural and biotechnological relevance. Microbes. Environ., 24(3): 205-216. Yu L, Kong F X, Zhang M, Yang Z, Shi X L, Du M Y. 2014. The dynamics of Microcystis genotypes and Microcystin production and associations with environmental factors during blooms in Lake Chaohu, China. Toxins, 6(12): 3 238-3 257.
Zan F Y, Huo S L, Xi B D, Li Q Q, Liao H Q, Zhang J T. 2011. Phosphorus distribution in the sediments of a shallow eutrophic lake, Lake Chaohu, China. Environ. Earth Sci., 62(8): 1 643-1 653.
Zan F Y, Huo S L, Xi B D, Zhu C W, Liao H Q, Zhang J T, Yeager K M. 2012. A 100-year sedimentary record of natural and anthropogenic impacts on a shallow eutrophic lake, Lake Chaohu, China. J. Environ. Monit., 14(3): 804-816.
Zhang L, Gao G, Tang X M, Shao K Q, Gong Y. 2016. Pyrosequencing analysis of bacterial communities in Lake Bosten, a large brackish inland lake in the arid northwest of China. Can. J. Microbiol., 62(6): 455-463.
Zhang L, Wang S R, Li Y P, Zhao H C, Qian W B. 2015. Spatial and temporal distributions of microorganisms and their role in the evolution of Erhai Lake eutrophication. Environ. Earth Sci., 74(5): 3 887-3 896.
Zhao Y, Wang Z Q, Yang Z P, Xie C P, Fan Q, Wang Y. 2002. Investigation on water pollution by algae at locations of water collection in Chaohu Lake. J. Environ. Health, 19(4): 316-318. (in Chinese with English abstract)
Zhou Q, Takenaka S, Murakami S, Seesuriyachan P, Kuntiya A, Aoki K. 2007. Screening and characterization of bacteria that can utilize ammonium and nitrate ions simultaneously under controlled cultural conditions. J. Biosci. Bioeng., 103(2): 185-191.
 Journal of Oceanology and Limnology2020年3期
Journal of Oceanology and Limnology2020年3期
- Journal of Oceanology and Limnology的其它文章
- List of the Most Outstanding Papers Published by CJOL/JOL in 2017-2018
- Who is the “murderer” of the bloom in coastal waters of Fujian, China, in 2019?*
- The investigation of internal solitary waves over a continental shelf-slope*
- Efference ect of diets on the feeding behavior and physiological properties of suspension-feeding sea cucumber Cucumaria frondosa*
- Efference ects of light quality on growth rates and pigments of Chaetoceros gracilis (Bacillariophyceae)*
- Marine bacterial surfactin CS30-2 induced necrosis-like cell death in Huh7.5 liver cancer cells*